(JO n° 291 du 2 décembre 2020)
NOR : EAEJ2029072D
Vus
Le Président de la République,
Sur le rapport du Premier ministre et du ministre de l'Europe et des affaires étrangères,
Vu la Constitution, notamment ses articles 52 à 55 ;
Vu le décret n° 53-192 du 14 mars 1953 modifié relatif à la ratification et à la publication des engagements internationaux souscrits par la France ;
Vu le décret n° 61-1300 du 30 novembre 1961 portant publication du traité sur l'Antarctique, signé le 1er décembre 1959 ;
Vu le décret n° 98-861 du 18 septembre 1998 portant publication du protocole au traité sur l'Antarctique, relatif à la protection de l'environnement, signé à Madrid le 4 octobre 1991 ;
Vu le décret n° 2005-1075 du 23 août 2005 portant publication de l'annexe V au protocole au traité de l'Antarctique relatif à la protection de l'environnement, protection et gestion des zones, adoptée à Bonn le 18 octobre 1991,
Décrète :
Article 1er du décret du 20 novembre 2020
Les mesures 1 à 6 (2018) relatives aux zones spécialement protégées de l'Antarctique, adoptées à Buenos Aires le 18 mai 2018, lors de la XLIe réunion consultative du traité sur l'Antarctique (RCTA), seront publiées au Journal officiel de la République française :
- Mesure 1 (2018) Zone spécialement protégée de l'Antarctique n° 108 (île Green, îles Berthelot, péninsule antarctique) : plan de gestion révisé (ensemble une annexe) ;
- Mesure 2 (2018) Zone spécialement protégée de l'Antarctique n° 117 (île Avian, baie Marguerite, péninsule antarctique) : plan de gestion révisé (ensemble une annexe) ;
- Mesure 3 (2018) Zone spécialement protégée de l'Antarctique n° 132 (péninsule Potter, île du Roi-George [Isla 25 de Mayo], îles Shetland du Sud) : plan de gestion révisé (ensemble une annexe) ;
- Mesure 4 (2018) Zone spécialement protégée de l'Antarctique n° 147 (vallée Ablation et mont Ganymède, île Alexandre) : plan de gestion révisé (ensemble une annexe) ;
- Mesure 5 (2018) Zone spécialement protégée de l'Antarctique n° 170 (Nunataks Marion, île Charcot, péninsule antarctique) : plan de gestion révisé (ensemble une annexe) ;
- Mesure 6 (2018) Zone spécialement protégée de l'Antarctique n° 172 (partie inférieure du glacier Taylor et Blood Falls de la vallée Taylor, dans les vallées sèches de McMurdo en terre Victoria) : plan de gestion révisé (ensemble une annexe).
Article 2 du décret du 20 novembre 2020
Le Premier ministre et le ministre de l'Europe et des affaires étrangères sont chargés, chacun en ce qui le concerne, de l'exécution du présent décret, qui sera publié au Journal officiel de la République française.
Fait le 30 novembre 2020.
Emmanuel Macron
Par le Président de la République :
Le Premier ministre,
Jean Castex
Le ministre de l'Europe et des affaires étrangères,
Jean-Yves Le Drian
Annexe
MESURE 1 (2018)
ZONE SPÉCIALEMENT PROTÉGÉE DE L'ANTARCTIQUE N° 108 (ILE GREEN, ILES BERTHELOT, PÉNINSULE ANTARCTIQUE) : PLAN DE GESTION RÉVISÉ (ENSEMBLE UNE ANNEXE)
Les Représentants,
Rappelant les articles 3, 5 et 6 de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement qui disposent de la désignation des zones spécialement protégées de l'Antarctique (« ZSPA ») et de l'adoption des plans de gestion pour ces zones ;
Rappelant :
- la recommandation IV-9 (1966) qui a désigné l'île Green, îles Berthelot, péninsule antarctique comme zone spécialement protégée (« ZSP ») n° 9 ;
- la recommandation XVI-6 (1991), qui a mis en annexe le plan de gestion de la zone ;
- la décision 1 (2002) qui a renommé et renuméroté la ZSP n° 9 en ZSPA nº 108 ;
- la mesure 1 (2002) et la mesure 1 (2013), qui ont adopté des plans de gestion révisés pour la ZSPA n° 108 ;
Rappelant que la résolution IV-9 (1966) a été désignée comme caduque par la décision 1 (2011) et que la recommandation XVI-6 (1991) n'est pas entrée en vigueur et a été retirée par la mesure 3 (2017) ;
Notant que le Comité pour la protection de l'environnement a approuvé un plan de gestion révisé pour la ZSPA nº 108 ;
Souhaitant remplacer le plan de gestion actuel pour la ZSPA nº 108 par le plan de gestion révisé ;
Recommandent à leurs gouvernements d'approuver la mesure suivante, conformément au paragraphe 1 de l'article 6 de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement :
Que :
1. Le plan de gestion révisé pour la zone spécialement protégée de l'Antarctique n° 108 (île Green, îles Berthelot, péninsule antarctique), qui figure en annexe de la présente mesure, soit approuvé ; et que
2. Le plan de gestion pour la zone spécialement protégée de l'Antarctique n° 108 figurant en annexe de mesure 1 (2013) soit abrogé.
Annexe
PLAN DE GESTION POUR LA ZONE SPÉCIALEMENT PROTÉGÉE DE L'ANTARCTIQUE NO 108
ÎLE GREEN, ÎLES BERTHELOT, PENINSULE ANTARCTIQUE
Introduction
La raison principale de la désignation de l'île Green, îles Berthelot, péninsule antarctique (65°19' de latitude sud, 64°09' de longitude ouest, 0,2 km2) comme zone spécialement protégée de l'Antarctique (ZSPA) est de protéger les valeurs environnementales et particulièrement celles de la tourbe tapissée de mousse Chorisodontium-Polytrichum présente dans la zone.
L'île Green avait été initialement désignée comme zone spécialement protégée (ZSP) en vertu de la recommandation IV-9 (1966, ZSP n° 9), suite à la proposition du Royaume-Uni. Cette désignation était justifiée par le fait que la végétation y « est exceptionnellement riche, [et] est probablement la plus luxuriante de la région occidentale de la péninsule antarctique ». La recommandation indiquait que : « à certains endroits l'épaisseur de l'humus atteignait 2 mètres et que cette zone, de par sa valeur scientifique, devait être protégée car elle était probablement l'un des écosystèmes les plus variés de l'Antarctique ». Un plan de gestion pour la zone a été préparé par le Royaume-Uni et adopté en vertu de la recommandation XVI-6 (1991). Les motifs de désignation avaient été étendus et précisés bien que la comparaison avec d'autres sites alentour donnait à penser que la flore de l'île Green n'était pas d'une diversité extraordinaire. Néanmoins, la flore de l'île avait été jugée digne d'intérêt notamment le versant nord de l'île, caractérisé par des bancs de tourbe couverte de mousse abondante et pérenne constituée de Chorisodontium aciphyllum et de Polytrichum strictum recouvrant essentiellement une tourbière de plus d'un mètre de profondeur. La canche antarctique (Deschampsia antarctica), l'une des deux seules plantes vasculaires poussant dans tout le territoire visé par le Traité sur l'Antarctique, avait été également observée en touffe parsemées sur les rochers abritant une colonie de cormorans impériaux (Phalacrocorax [atriceps] bransfieldensis). Cette colonie de cormorans, que l'on rencontre dans les hauteurs abruptes au nord-ouest de l'île, avait été identifiée comme une des plus importantes de la péninsule antarctique. Le plan de gestion avait été révisé par le biais de la Mesure 1 (2002).
Cet espace s'inscrit bien dans le cadre du système plus global de désignation de zones spécialement protégées puisqu'il abrite une tourbe couverte de mousse ainsi qu'une tourbière présentant des caractéristiques rares dans l'ouest de la Péninsule antarctique. Ces bancs de mousses se distinguent de celles des ZSPA plus au nord car elles sont très peu touchées par l'impact nuisible de l'otarie de Kerguelen (Arctocephalus gazella). La résolution 3 (2008) recommandait que l'analyse des domaines environnementaux pour le continent antarctique serve de modèle dynamique pour l'identification des zones spécialement protégées de l'Antarctique dans le cadre environnemental et géographique systématisé visé à l'article 3 (2) de l'annexe V du Protocole (voir également Morgan et al., 2007). Selon ce modèle, la ZSPA n° 108 relève du domaine environnemental B (géologie des latitudes septentrionales moyennes de la péninsule antarctique). Parmi les autres zones protégées contenant le Domaine B, on compte notamment les ZSPA 115, 134, 140 et 153 et la ZGSA n° 4. La ZSPA n° 108 se trouve dans la région de conservation biogéographique de l'Antarctique (RCBA) 3 : nord-ouest de la péninsule antarctique.
1. Description des valeurs à protéger
A la suite d'une visite de gestion de la ZSPA en avril 2017, les valeurs énoncées dans la désignation antérieure ont été réaffirmées. Ces valeurs sont exposées comme suit :
Les bancs de mousse Polytrichum strictum et de Chorisodontium-Polytrichum constituent la raison première de cette protection spéciale, car elles sont peut-être l'un des paysages les plus représentatifs de ce type de flore dans l'ouest de la péninsule antarctique, occupant une superficie de plus de 0,5 hectare.
En outre, au cours des dernières années, de nombreux bancs de mousse comparables situés sur des îles plus au nord ont subi les répercussions négatives de la croissance de la population d'otaries de Kerguelen.
La végétation de l'île Green a jusqu'à présent échappé à tout dégât majeur.
La mousse Chorisodontium aciphyllum est présente à la limite méridionale de l'île vers les îles Berthelot.
La zone abrite une importante colonie de cormorans impériaux (Phalacrocorax [atriceps] bransfieldensis), qui est probablement l'une des espèces qui se reproduisent le plus dans la péninsule antarctique.
L'île Green a bénéficié d'une protection pendant pratiquement toutes les périodes d'activités scientifiques dans la région et des autorisations d'accès ont été délivrées uniquement pour mener à bien des recherches scientifiques indispensables. L'île n'ayant pas subi de visites, de recherches, ni de prélèvements d'échantillons intensifs, elle pourrait constituer un site de référence intéressant pour de futures études.
2. Buts et objectifs
Le plan de gestion destiné à l'île Green vise à :
- éviter toute détérioration ou tout risque de détérioration des valeurs de la zone en empêchant toute perturbation humaine inutile de ladite zone ;
- éviter ou réduire au maximum l'introduction de plantes, d'animaux et de microbes non indigènes dans la zone ;
- limiter le risque d'introduction d'agents pathogènes qui pourraient provoquer des maladies au sein des populations fauniques de la zone ;
- permettre d'effectuer des recherches scientifiques dans la zone, pour autant qu'elles soient indispensables, qu'elles ne puissent être menées ailleurs et qu'elles ne portent pas atteinte à l'écosystème naturel de la zone ; et
- préserver l'écosystème naturel de la zone pour servir ultérieurement de zone de référence dans de futures études.
3. Activités de gestion
Les activités de gestion suivantes devront être menées à bien pour protéger les valeurs de la zone :
- des copies de ce plan de gestion doivent être mises à la disposition des navires et aéronefs ayant l'intention de visiter les alentours de la zone ;
- les bornes, les panneaux ou autres structures (par ex. cairns) érigés dans la zone à des fins scientifiques et de gestion seront sécurisés et maintenus en bon état, puis enlevés lors qu'ils ne seront plus nécessaires ;
- le plan de gestion doit être réexaminé au moins une fois tous les cinq ans et mis à jour en conséquence ;
- une copie de ce plan de gestion sera mise à disposition à la station Akademik Vernadsky (Ukraine ; 65°15' de latitude sud, 64°16' de latitude ouest) ;
- toutes les activités scientifiques et de gestion entreprises au sein de la zone devraient faire l'objet d'une évaluation d'impact sur l'environnement, conformément à ce que requiert l'annexe I du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement ;
- les directeurs des programmes antarctiques nationaux en cours d'exécution dans la région se livreront entre eux à des consultations pour veiller à ce que les activités de gestion susmentionnées soient mises en œuvre.
4. Durée de la désignation
La zone est désignée pour une période indéterminée.
5. Cartes et photographies
Carte 1. - Carte de synthèse, montrant l'emplacement de l'île Green sur la péninsule antarctique. spécifications de la carte : WGS84 Stéréographique polaire antarctique. Méridien central - 55°, parallèle de référence : - 71°.
Carte 2. - Carte de la région montrant l'emplacement de la ZSPA n° 108, île Green, îles Berthelot, par rapport aux stations et aux autres zones protégées alentours. Spécifications de la carte : WGS84 Stéréographique polaire antarctique. Méridien central -64°, parallèle de référence : - 71°.
Carte 3. - ZSPA n° 108, île Green, îles Berthelot, péninsule antarctique, carte topographique. carte réalisée à partir d'une étude de terrain le 24 février 2001 et d'une orthophotographie numérique (photographie aérienne de la British Antarctic Survey, prise le 14 février 2001). Spécifications de la carte - Projection : zone UTM 20S ; sphéroïde : WGS84 ; Datum : niveau moyen de la mer (EGM96).
6. Description de la zone
6 (i) Coordonnées géographiques, bornage et caractéristiques milieu naturel
Description générale
L'île Green (65°19' de latitude sud, 64°09' de latitude ouest, environ 0,2 km2 ; carte 1) est une petite île située à 150 mètres au nord de la plus grande des îles Berthelot, canal Grandidier, à environ 3 km au large de la côte Graham sur la péninsule antarctique (carte 2). Elle s'étend sur 520 mètres du nord au sud sur 500 mètres d'est en ouest. Le relief culmine en un pic arrondi à 83 mètres d'altitude. L'île est entourée de flancs raides habillés de falaises vertigineuses au sud et à l'est. Les terres basses de l'île sont essentiellement situées sur la côte septentrionale, où les rochers descendent en pente douce. Des neiges éternelles sont présentes, et surtout autour du sommet ainsi que l'espace situé en altitude au sud et à l'est du point culminant. Il n'existe aucun plan d'eau douce permanent sur l'île.
Limites
La zone désignée comprend la totalité de l'île Green, ses limites étant définies en fonction du niveau de la mer à marée basse. Les îlots et rochers alentours ne sont pas inclus dans la zone. Aucune borne n'a été installée. La côte est très clairement définie et les limites de la zone sont visibles à l'œil nu.
Climat
Il n'existe pas de données d'archive détaillées sur la météorologie de l'île Green, mais les conditions doivent être semblables à celles de la station Akademik Vernadsky (Ukraine) sur l'île Galindez, îles Argentine, située à 8 km au nord. La température estivale moyenne à la station est de 0°C, tandis que la température maximale est de 11,7°C. En hiver, la température moyenne est de -10°C, le mercure peut descendre jusqu'à -43,3°C. La vitesse moyenne du vent est de 7,5 nœuds.
Géologie et sols
L'île Green, à l'instar du reste des îles Berthelot, est composée de gabbro du Jurassique inférieur - Tertiaire inférieur (British Antarctic Survey, 1981). Hormis les importants dépôts de tourbe, le sol dépasse rarement les 20 cm de profondeur sauf dans les ravines et les dépressions rocheuses. Il est principalement constitué de minéraux à humiques grossiers issus de l'altération de la roche-mère. Les saillies rocheuses et les ravines situées à proximité de la colonie de cormorans impériaux sont constituées d'un sol plus riche, dérivé en partie de mousses et de guano décomposés. Sur la majeure partie du relief en pentes raides au nord de l'île, les mousses Chorisodontium aciphyllum et Polytrichum strictum ont développé un tapis épais de mousse vivante recouvrant au moins un mètre de tourbière à peine altérée ou décomposée (Smith, 1979, Fenton et Smith, 1982). L'étude des propriétés de la tourbière pourrait contribuer à l'identification des caractéristiques climatiques de l'Holocène tardif (Royles et al. 2012). La couche de pergélisol se trouve à une profondeur de 20 à 30 cm sous le sol. Ailleurs sur l'île, notamment sur le flanc nord-est, des petites zones d'éboulis ont été observées. Aucune particularité périglaciaire bien développée n'a pu être constatée malgré la présence évidente de quelques cercles de pierre.
Végétation
Le Polytrichum strictum est la variété qui domine incontestablement la flore de l'île, présente sur les reliefs en pente au nord de l'île (carte 3). La zone de végétation, d'une largeur de près de 140 mètres, s'étend à une altitude variant entre 25 et 70 mètres et couvre une étendue de plus de 0,5 ha (Bonner et Smith, 1985). Les techniques de télédétection par satellite (Indice différentiel normalisé de végétation) ont révélé que la surface de végétation verte dans la ZSPA était de 0,036 km2 (environ 16,5 % de la surface de la ZSPA). La végétation de la zone est luxuriante et la tourbe gelée peut atteindre deux mètres de profondeur. La surface de la mousse sèche et compacte est étagée, ce qui serait le résultat de la descente de couches de tourbe active vers les flancs abruptes des falaises. L'érosion des bancs de mousse est très marquée à certains endroits, ce qui semble dû au fait que la tourbière atteint une profondeur critique et déborde sur les pentes. Les otaries ne seraient pas en cause dans cette évolution, contrairement aux résultats des observations faites sur des ZSPA situées plus au nord (p. ex. : ZSPA n° 113). La variété Chorisodontium aciphyllum est plus répandue sur les bordures des bancs de mousse et autour des petites ravines qui s'y forment. En effet, l'écosystème offre une protection et l'humidité provenant de la congère lui est favorable. Ces deux genres de mousses sont souvent amalgamés dans les grandes tourbières du nord de l'Antarctique maritime. Cependant dans la région du canal Grandidier, on retrouve une mousse plus xérique, exclusivement constituée de P. strictum. La C. aciphyllum est plus répandue à la limite méridionale de l'île Green (Smith, 1996). On retrouve fréquemment les variantes de C. aciphyllum telles que Pohlia nutans et les hépatiques Barbilophozia hatcheri et Cephaloziella varians. Les lichens épiphytes ne croissent pas souvent sur la mousse vivante constituée de Polytrichum et de Chorisodontium, mais le Sphaerophorus globosus peut être identifié dans les espaces plus exposées du nord-ouest. Plusieurs espèces de Cladonia se retrouvent souvent sur les bancs de mousse. L'Ochrolechia frigida, lichen épiphyte blanc et verruqueux, peut également se retrouver dans une moindre mesure dans cette zone. Les espèces de mousses noires encroûtâtes s'épanouissent généralement dans la mousse moribonde.
Les petites étendues de mousse constituées de Warnstorfia fontinaliopsis, Brachythecium austro-salebrosum et Sanionia uncinata poussent généralement dans les cavités humides entre les roches et dans les galeries de fonte. Partout ailleurs, la végétation est dominée par les lichens. On peut observer la présence dominante des communautés de Usnea antarctica et de Umbilicaria (notamment les genres U. antarctica, U. decussate, U. hyperborea et U. umbilicarioides) qui poussent sur les rochers et les galets loin de la côte et des oiseaux marins. Il est toutefois possible d'identifier dans cet espace les mousses Andreaea depressinervis et A. regularis ainsi que plusieurs genres de lichens encroûtants. Les falaises surplombant le littoral abritent les communautés les plus diversifiées et les plus hétérogènes. Elles sont essentiellement composées de lichens issus de la modification de la communauté de Usnea-Umbilicaria au contact de plusieurs taxons « nitrophiles » (dépendant de l'azote) présents à proximité des nids d'oiseaux marins, notamment les Acarospora, Buellia, Caloplaca, Lecanora, Mastodia, Omphalodina, Physcia et Xanthoria. Les recensements de plantes dans la région ont servi de référence pour la réalisation de projections régionales et locales concernant la diversité de la flore de lichen dans la péninsule antarctique (Casanovas et al. 2012). La seule plante à fleurs répertoriée à ce jour sur l'île Green est la canche antarctique (Deschampsia antarctica), qui pousse fréquemment en petites touffes parsemées au-dessus des sites abritant les colonies de cormorans et sur les saillies rocheuses à l'ouest de l'île. Quant aux algues vertes foliacées Prasiola crispa, elles sont très répandues dans les espaces humides de l'île.
Oiseaux nicheurs
Une importante colonie de cormorans impériaux (Phalacrocorax atriceps) évolue sur un flanc raide et rocheux au nord-ouest de l'île (65°19'21''latitude sud, 64°09'11'' longitude ouest ; carte 3). Il s'agit de l'une des plus grandes colonies de cormorans impériaux identifiées dans la péninsule antarctique (Bonner et Smith, 1985), quoique les effectifs varient considérablement d'une année à l'autre (Casaux et Barrera-Oro, 2006). En 1971, on dénombrait dans la zone 50 couples environ (Kinnear, 1971). En 1973, on comptait 112 individus (Schlatter et Moreno, 1976). Lors d'une visite réalisée en mars 1981, 500-600 individus avaient été recensés (dont 300-400 juvéniles). Le 24 février 2001, Harris (Harris, 2001) avait dénombré 71 oisillons. Le recensement effectué le 15 février 2011 a permis d'identifier 100 individus. Le 22 janvier 2013, l'effectif était de 200-250 individus dont 100 adultes. En avril 2017, environ 100 adultes ont été observés. Les labbes bruns (Stercorarius antarcticus) sont très répandus dans l'île, ils occupent généralement les grands bancs de mousse. Le labbe de McCormick (Stercorarius maccormicki) fait également partie de la faune de l'île. Quelques spécimens hybrides issus de ces oiseaux sont parfois représentés. Plus de 80 oiseaux avaient été observés en mars 1981 dont 10 couples nicheurs, élevant essentiellement des nichées de deux oisillons. La visite n'avait pas permis d'identifier d'autres oiseaux.
Invertébrés
Les données concernant les invertébrés présents sur l'île Green sont assez limitées. Cependant 15 espèces d'invertébrés avaient été recensées dans le cadre d'une étude indiquant que la faune d'invertébrés était assez diversifiée pour la région (Usher et Edwards, 1986). Les espèces les plus représentées étaient : Cryptopygus antarcticus, Belgica antarctica et Nanorchestes gressitti. La larve B.antarctica était beaucoup plus répandue sur l'île Green que sur l'île voisine : l'île Darboux Les autres espèces rencontrées dans la zone sont les suivantes : Alaskozetes antarcticus, Ereynetes macquariensis, Eupodes minutus, Eupodes parvus grahamensis, Friesea grisea, Gamasellus racovitzai, Halozetes belgicae, N. berryi, Oppia loxolineata, Parisotoma octo-oculata, Rhagidia gerlachei et Stereotydeus villosus.
Activités humaines et leur impact
Les visites de l'île Green qui ont été documentées sont assez rares. Le premier débarquement sur l'île dont la mention est connue date de la Première expédition antarctique française en 1903-1905. Puis il y eut quelques autres visites par la Deuxième expédition antarctique française au cours de l'hiver 1909. La British Graham Land Expedition débarque sur l'île le 18 mars 1935. Des études sur la flore de l'île Green ont été entreprises par Smith en 1981 (Bonner et Smith, 1985) et Komárková en 1982-1983 (Komárková, 1983). En janvier 1989, une équipe d'inspection (Heap, 1994) a enregistré (et laissé sur place) de nombreux bouts de fil de fer d'environ 30 cm de long et 2,5 mm de diamètre, délimitant les angles d'une aire de 50 m2 : il s'agissait d'une tourbière recouverte d'un tapis de mousse Polytrichum strictum. Il n'a pas été possible de déterminer avec précision la période à laquelle ces repères ont été installés. Le nombre de repères, leurs emplacements ainsi que les caractéristiques d'une éventuelle pollution de la mousse par ce matériel sont inconnus. En janvier 2013, une tringle métallique d'environ 20 cm de long, dont l'origine est inconnue, a été retrouvée sur la mousse à 65°19'23"de latitude sud et 64°09'02''de longitude ouest.
Plus récemment, la flore de plusieurs sites situés dans la péninsule antarctique a été endommagée par le piétinement et l'enrichissement excessif de l'eau de mer par des nutriments, dus à la présence de l'otarie de Kerguelen (Arctocephalus gazella). La présence d'otarie de Kerguelen n'a pas été détectée sur l'île Green lors de la visite du 24 février 2001, bien que des traces récentes de piétinement et des indices d'enrichissement des sols aient été observés à plusieurs endroits sur la partie inférieure des bancs de mousse. Cependant, les dommages causés au site étaient très limités et la plupart des bancs de mousses étaient bien préservés. Une visite effectuée en avril 2017 n'a pas révélé de nouvelles perturbations par les otaries.
6 (ii) Accès à la zone
L'accès à la zone est autorisé aux embarcations, ou aux véhicules et aux piétons en empruntant la banquise. Il n y a pas de restriction quant à l'itinéraire d'accès à la zone, ou pour la quitter, par embarcation ou par la banquise.
Le site de débarquement des petites embarcations se trouvera de préférence sur la côte rocheuse septentrionale. L'aire de débarquement recommandée est située sur une petite crique à 65°19'17,6'' de latitude sud et 64°08'46,0'' de longitude ouest (carte 3). L'accès en petite embarcation à d'autres endroits autour de la côte est autorisé pour autant qu'il soit conforme aux objectifs pour lesquels le permis a été délivré.
Lorsque l'accès par la glace de mer est possible, il n'existe aucune restriction quant aux endroits où les véhicules ou les piétons peuvent accéder au site, étant entendu que les véhicules ne peuvent en aucun cas être utilisés sur la terre ferme.
L'atterrissage d'aéronefs dans la zone est interdit.
Les équipages et autres personnes à bord des embarcations ne sont pas autorisés à se déplacer à pied dans les alentours immédiats du site de débarquement, sauf autorisation expresse prévue par le permis.
6 (iii) Structures à l'intérieur et à proximité de la zone
Aucune structure n'est installée dans la zone. La station de recherche scientifique la plus proche est Akademik Vernadsky (Ukraine) (65°15'de latitude sud, 64°16'de longitude ouest). Elle se trouve à environ 8 km au nord de la zone, sur l'île Galindez.
6 (iv) Emplacement d'autres zones protégées à proximité
Les autres zones protégées dans les environs sont :
- ZSPA n° 113, île Lichfield, port Arthur, île Anvers, archipel Palmer, 64°46' de latitude sud, 64°06' de longitude ouest, 62 km au nord ;
- ZSPA n° 139, pointe Biscoe, île Anvers, archipel Palmer, 64°48' de latitude sud, 63°46' de longitude ouest, 60 km au nord ;
- ZSPA n° 146, baie du Sud, île Doumer, archipel Palmer, 64°51' de latitude sud, 63°34' longitude ouest, 60 km au nord-ouest.
Les ZSPA n° 113 et 139 sont situées dans la zone gérée spéciale de l'Antarctique n° 7, île Southwest Anvers et bassin Palmer.
6 (v) zones spéciales à l'intérieur de la zone
Il n'y a aucune zone spéciale à l'intérieur de la zone.
7. Critères de délivrance des permis
7 (i) Conditions générales pour l'obtention d'un permis
L'accès à la zone est interdit sauf avec un permis délivré par une autorité nationale compétente. Les critères de délivrance d'un permis pour entrer dans la zone sont les suivants :
- un permis est délivré pour des raisons scientifiques indispensables qu'il est impossible de satisfaire ailleurs ou pour des raisons de gestion essentielles à la zone ;
- les activités autorisées sont conformes au présent plan de gestion ;
- toutes les activités de gestion soutiennent la réalisation des buts et objectifs du présent plan de gestion ;
- les actions autorisées ne mettront pas en péril l'écosystème naturel de la zone ;
- les activités autorisées veilleront, au moyen d'un processus d'évaluation d'impact sur l'environnement, à la protection permanente des valeurs environnementales ou scientifiques de la zone ;
- le permis est délivré pour une durée déterminée ; et
- le permis, ou une copie certifiée, sera emporté à l'intérieur de la zone.
7 (ii) Accès à la zone et déplacements à l'intérieur de celle-ci
L'accès à la zone est interdit aux véhicules terrestres et les déplacements doivent se faire exclusivement à pied.
Les opérations de survol de la zone doivent être réalisées conformément aux Directives pour l'exploitation d'aéronefs à proximité de concentrations d'oiseaux dans l'Antarctique, inscrites dans la résolution 2 (2004).
Le survol de colonies d'oiseaux dans la zone par des systèmes d'aéronefs pilotés à distance (RPAS) n'est pas autorisé, sauf à des fins scientifiques ou opérationnelles, et en vertu d'un permis émis par une autorité nationale compétente.
Les piétons doivent prendre toutes les précautions utiles pour minimiser l'impact de leur présence sur le sol, la couverture végétale et les oiseaux, notamment en marchant sur les surfaces enneigées ou les terrains rocailleux.
Les déplacements à pied doivent être limités au minimum requis pour effectuer les activités autorisées et tous les efforts raisonnables doivent être consentis pour réduire les effets du piétinement.
7 (iii) Activités pouvant être conduites à l'intérieur de la zone
Les activités pouvant être menées sont :
- les activités de gestion essentielles, y compris de suivi ;
- des travaux de recherche scientifique indispensables qui ne peuvent être entrepris ailleurs et ne risquent pas de mettre en péril l'écosystème de la zone ; et
- l'échantillonnage qui doit être réduit au minimum pour répondre aux programmes de recherches approuvés.
7 (iv) Installation, modification ou enlèvement de structures
Les structures ou installations permanentes sont interdites.
Aucune structure ne doit être érigée dans la zone et aucun matériel scientifique ne doit y être installé, sauf pour des raisons scientifiques ou de gestion indispensables et pour une période préétablie définies dans un permis.
Toutes les bornes, les structures et tout l'équipement scientifique installés dans la zone doivent clairement identifier le pays, le nom du principal chercheur ou de la principale agence, l'année d'installation et la date d'enlèvement prévue.
Tous ces objets ne doivent contenir aucun organisme, propagule (par ex., semence, œufs, spores) ou terre non stérile (voir section 7 [vi]), et doivent être composés de matériaux résistants aux conditions environnementales et présenter un risque de contamination minime pour la zone.
L'enlèvement d'équipements spécifiques pour lesquels le permis a expiré est du ressort de l'autorité qui a délivré le permis d'origine et il sera l'un des critères régissant la délivrance du permis.
7 (v) Emplacement des camps
Lorsqu'ils sont indispensables pour remplir des objectifs entrant dans le cadre d'un permis, les campements temporaires sont autorisés dans la zone, au niveau de la plateforme située sur la côte septentrionale (65°19'18'' de latitude sud, 64°08'55'' de longitude ouest ; carte 3). Les camps doivent être installés sur les surfaces enneigées stables qui caractérisent cet emplacement, ou sur le gravier ou les rochers en l'absence de couverture neigeuse. L'installation de camps sur les zones ayant une couverture végétale permanente est interdite.
7 (vi) Restrictions relatives aux matériaux et organismes pouvant être introduits dans la zone
L'introduction délibérée dans la zone d'animaux, de végétaux ou de micro-organismes vivants est interdite. Pour garantir le maintien des valeurs floristiques et écologiques de la zone, des précautions spéciales devront être prises pour prévenir toute introduction accidentelle de microbes, d'invertébrés ou de plantes provenant d'autres sites antarctiques, y compris des stations, ou d'autres régions que l'Antarctique. Tout le matériel d'échantillonnage et les balises introduits dans la zone doivent être nettoyés et stérilisés. Dans la mesure du possible, les chaussures et autres équipements utilisés ou introduits dans la zone (y compris les sacs et les sacs à dos) doivent être minutieusement nettoyés avant d'entrer dans la zone. Le Manuel sur les espèces non-indigènes du CPE (édition 2017) et le Environmental code of conduct for terrestrial scientific field research in Antarctica [Code de conduite environnementale pour la recherche scientifique sur le terrain en antarctique] (SCAR 2009). Compte tenu de la présence de colonies d'oiseaux nicheurs dans la zone, aucun produit provenant ou dérivé d'espèces avicoles - notamment les déchets, les produits contenant des œufs en poudre non pasteurisés - ne doit être introduit dans la zone ou déversé dans la mer au large ou à proximité de la zone.
Aucun herbicide ni pesticide ne doit être introduit dans la zone. Tout autre produit chimique, y compris les radionucléides ou les isotopes stables, qui peuvent être introduits pour des raisons scientifiques ou des raisons de gestion visées dans le permis, seront enlevés de la zone au plus tard à la fin de l'activité pour laquelle le permis a été délivré. L'émission, directement dans l'environnement, de radionucléides ou d'isotopes stables par une méthode les rendant irrécupérables doit être évitée. Les carburants et autres produits chimiques ne doivent pas être entreposés dans la zone, hormis sur délivrance d'un permis. Auquel cas, ces matériaux doivent être entreposés et manipulés de façon à limiter les risques d'introduction accidentelle dans l'environnement. Tous les matériaux sont introduits dans la zone pour une période déterminée uniquement ; ils doivent être enlevés lorsque cette période est échue. En cas de déversement susceptible de mettre en péril les valeurs de la zone, leur retrait est encouragé à condition que l'impact de celui-ci ne soit pas susceptible d'être supérieur à celui consistant à laisser les substances in situ. L'autorité compétente doit être notifiée de tout élément introduit et non retiré qui ne figurait pas dans le permis agréé.
7 (vii) Prélèvement de végétaux, capture d'animaux ou perturbations nuisibles de la faune et la flore
Toute capture d'animaux ou toute perturbation nuisible à la faune et la flore indigène est interdite sauf avec un permis distinct délivré spécifiquement à cette fin en vertu de l'annexe II du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement. Dans le cas de prélèvements ou de perturbations nuisibles d'animaux, le SCAR code of conduct for the use of animals for scientific purposes in Antarctica [Code de conduite du SCAR pour l'utilisation d'animaux à des fins scientifiques dans l'Antarctique] (2011) devra être utilisé comme norme minimale. Tout prélèvement du sol ou de la flore à des fins d'échantillonnage doit se limiter au strict minimum nécessaire aux activités scientifiques et de gestion. Les techniques envisagées à cet effet doivent avoir les moindres répercussions possibles sur le sol, la glace et le biote.
7 (viii) Collecte ou retrait de matériaux non introduits dans la zone par le titulaire du permis
Les matériaux ne peuvent être ramassés ou enlevés de la zone qu'en conformité avec un permis, mais ils doivent être limités au minimum requis pour répondre aux besoins scientifiques ou de gestion. Les matériaux d'origine humaine susceptibles de mettre en péril les valeurs de la zone, qui n'ont pas été introduits dans celle-ci par le détenteur du permis ou qui n'ont pas été autrement autorisés, peuvent être enlevés de la zone à moins que l'impact environnemental de l'enlèvement ne soit plus grand que si les matériaux sont laissés in situ. Si tel est le cas, l'autorité compétente doit en être informée et son autorisation obtenue.
7 (ix) Elimination des déchets
Tous les déchets, y compris les déchets humains, doivent être retirés de la zone. Les déchets d'origine humaine peuvent être jetés à la mer.
7 (x) Mesures qui peuvent être nécessaires pour continuer de répondre aux objectifs du plan de gestion
Des permis peuvent être délivrés pour entrer dans la zone afin d'y réaliser des travaux de recherche scientifique, de surveillance et d'inspection de site, qui font intervenir le prélèvement d'un petit nombre d'échantillons à des fins d'analyse, ou pour appliquer des mesures de protection.
Tous les sites de suivi de longue durée doivent être signalés par des bornes ou des panneaux dûment entretenus.
Les activités de nature scientifique seront menées conformément au Environmental code of conduct for terrestrial scientific field research in Antarctica [Code de conduite environnemental pour la recherche scientifique sur le terrain en Antarctique] (SCAR, 2009).
7 (xi) Rapports de visite
Le principal détenteur du permis soumettra, pour chaque visite dans la zone, un rapport à l'autorité nationale compétente, dès que possible, et au plus tard six mois après la fin de ladite visite. Ces rapports doivent contenir, le cas échéant, les catégories d'informations mentionnées dans le formulaire de rapport de visite repris dans le Guide révisé pour l'élaboration des plans de gestion pour les zones spécialement protégées de l'Antarctique (annexe 2). Les autorités compétentes doivent être informées de toute activité ou mesure qui ne serait pas autorisée par le permis. Dans la mesure du possible, l'autorité nationale doit également transmettre une copie du rapport de visite à la Partie qui a proposé le plan de gestion et ce, afin de contribuer à gérer la zone et à revoir le plan de gestion. Les Parties doivent, dans la mesure du possible, déposer les originaux ou les copies de ces rapports dans une archive à laquelle le public pourra avoir accès afin de conserver une archive d'usage qui sera utilisée pour toute révision du plan de gestion et pour l'organisation de l'utilisation scientifique de la zone.
8. Bibliographie
Bonner, W. N., et Smith, R. I. L. (Eds.). (1985). Conservation areas in the Antarctic. SCAR, Cambridge : 73- 84.
Booth, R. G., Edwards, M. , et Usher, M. B. (1985). Mites of the genus Eupodes (Acari, Prostigmata) from maritime Antarctica : a biometrical and taxonomic study. Journal of Zoology 207 : 381-406.
British Antarctic Survey. (1981). carte géologique (Echelle 1 : 500000). Series BAS 500G Sheet 3, Edition 1.
Cambridge : British Antarctic Survey.
Casanovas, P., Lynch, H. L., et Fagan, W. F. (2012). Multi-scale patterns of moss and lichen richness on the Antarctic Peninsula. Ecography 35 : 001-011.
Casaux, R., et Barrera-Oro, E. (2006). Review. Shags in Antarctica : their feeding behaviour and ecological role in the marine food web. Antarctic Science 18 : 3-14.
Comité pour la protection de l'environnement (CPE). (2017). Manuel sur les espèces non indigènes - 2e édition. Manuel préparé par le Groupe de contact intersessions du CPE et adopté par la Réunion consultative du Traité sur l'Antarctique par la résolution 4 (2017). Buenos Aires, Secrétariat du Traité sur l'Antarctique.
Corner, R. W. M. (1964). Biological report (interim) for Argentine Islands. Rapport non publié, British Antarctic Survey Archives Ref AD6/2F/1964/N1.
Fenton, J. H. C, et Smith, R. I. L. (1982). Distribution, composition and general characteristics of the moss banks of the maritime Antarctic. British Antarctic Survey Bulletin 51 : 215-236.
Greene, D. M, et Holtom, A. (1971). Studies in Colobanthus quitensis (Kunth) Bartl. and Deschampsia antarctica Desv. : III. Distribution, habitats and performance in the Antarctic botanical zone. British Antarctic Survey Bulletin 26 : 1-29.
Harris, C. M. (2001). Revision of management plans for Antarctic protected areas originally proposed by the United States of America and the United Kingdom : Field visit report. Internal report for the National Science Foundation, US, and the Foreign and Commonwealth Office, UK. Environmental Research and Assessment, Cambridge.
Heap, J. (Ed.). (1994). Manuel du Système du Traité sur l'Antarctique. 8 e édition. Département d'Etat américain, Washington.
Hughes, K. A., Ireland, L. C., Convey, P., et Fleming, A. H. (2016). Assessing the effectiveness of specially protected areas for conservation of Antarctica's botanical diversity. Conservation Biology, 30 : 113- 120.
Kinnear, P. K. (1971). Phalacrocorax atriceps population data cited in BAS internal report - original reference unavailable.
Komárková, V. (1983). Studies of plant communities of the Antarctic Peninsula near Palmer Station.
Antarctic Journal of the United States 18 : 216-218.
Royles, J., Ogée, J., Wingate, L., Hodgson, D. A., Convey, P., et Griffiths, H. (2012). Carbon isotope evidence for recent climate-related enhancement of CO2 assimilation and peat accumulation rates in Antarctica. Global Change Biology 18 : 3112-3124.
SCAR (Comité scientifique pour la recherche antarctique). (2009). Environmental code of conduct for terrestrial scientific field research in Antarctica [Code de conduite environnemental pour la recherche scientifique sur le terrain en Antarctique]. XXXIIe RCTA IP4.
SCAR (Comité scientifique pour la recherche antarctique). (2011). SCAR code of conduct for the use of animals for scientific purposes in Antarctica [Code de conduite du SCAR pour l'utilisation d'animaux à des fins scientifiques en Antarctique]. XXXIVe RCTA IP53.
Schlatter, R. P., et Moreno, C. A. (1976). Habitos alimentarios del cormoran Antartico, Phalacrocorax atriceps bransfieldensis (Murphy) en Isla Green, Antartica. Serie Cientificia, Instituto Antártico Chileno 4 (1) : 69-88.
Smith, M. J., et Holroyd, P. C. (1978). 1978 Travel report for Faraday. Rapport non publié, British Antarctic Survey Archives Ref AD6/2F/1978/K.
Smith, R. I. L. (1979). Peat forming vegetation in the Antarctic. Dans : Proceedings of the International Symposium on Classification of Peat and Peatlands Finland, September 17-21, 1979. International Peat Society : 58-67).
Smith, R. I. L. (1982). Farthest south and highest occurrences of vascular plants in the Antarctic. Polar Record 21 : 170-173.
Smith, R. I. L. (1996). Terrestrial and freshwater biotic components of the western Antarctic Peninsula.
Dans : Ross, R.M., Hofmann, E.E., et Quetin, L.B. (Eds.) Foundations for ecological research west
of the Antarctic Peninsula. Antarctic Research Series 70 : 15-59.
Smith, R. I. L., et Corner, R.W. M. (1973). Vegetation of Arthur Harbour - Argentine Islands Region.
British Antarctic Survey Bulletin 33&34 : 89-122.
Stark, P. (1994). Climatic warming in the central Antarctic Peninsula area. Weather 49 (6) : 215-220. Terauds, A., et Lee, J. R. (2016). Antarctic biogeography revisited : updating the Antarctic Conservation
Biogeographic Regions. Diversity and Distribution 22 : 836-840.
Terauds, A., Chown, S. L., Morgan, F., Peat, H. J., Watt, D., Keys, H., Convey, P., et Bergstrom, D. M. (2012). Conservation biogeography of the Antarctic. Diversity and Distributions 18 : 726-41.
Usher, M. B., et Edwards, M. (1986). The selection of conservation areas in Antarctica : an example using the arthropod fauna of Antarctic islands. Environmental Conservation 13 (2) : 115-122.
Carte 1. - Carte de synthèse, montrant l'emplacement de l'île Green sur la péninsule antarctique. Spécifications de la carte : WGS84 Stéréographique polaire antarctique. Méridien central -55°, parallèle de référence : -71.
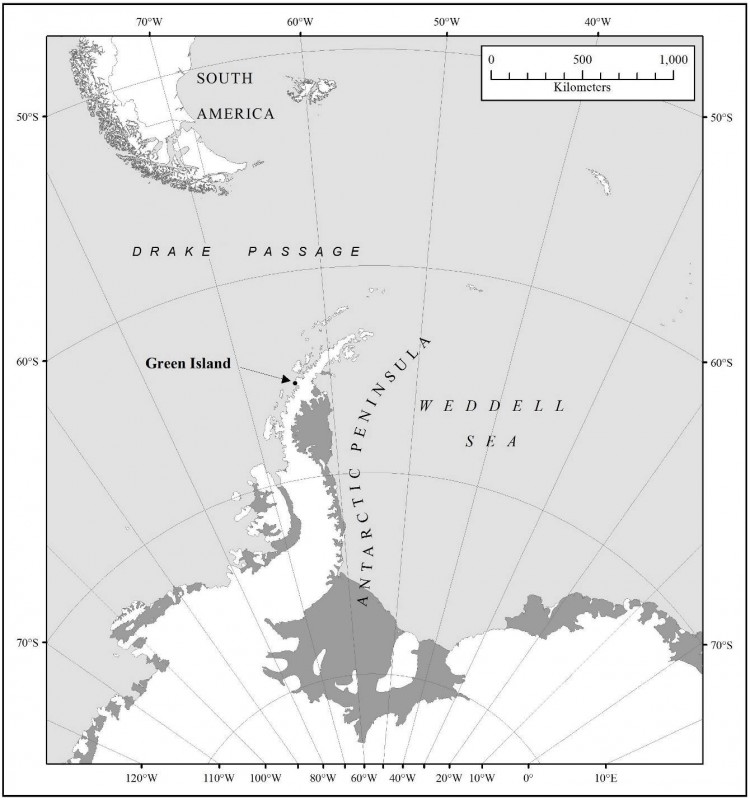
Carte 2. - Carte de la région montrant l'emplacement de la ZSPA n° 108, île Green, îles Berthelot, par rapport aux stations et aux autres zones protégées alentours. Spécifications de la carte : WGS84 Stéréographique polaire antarctique. Méridien central -64 o, parallèle de référence : -71°.
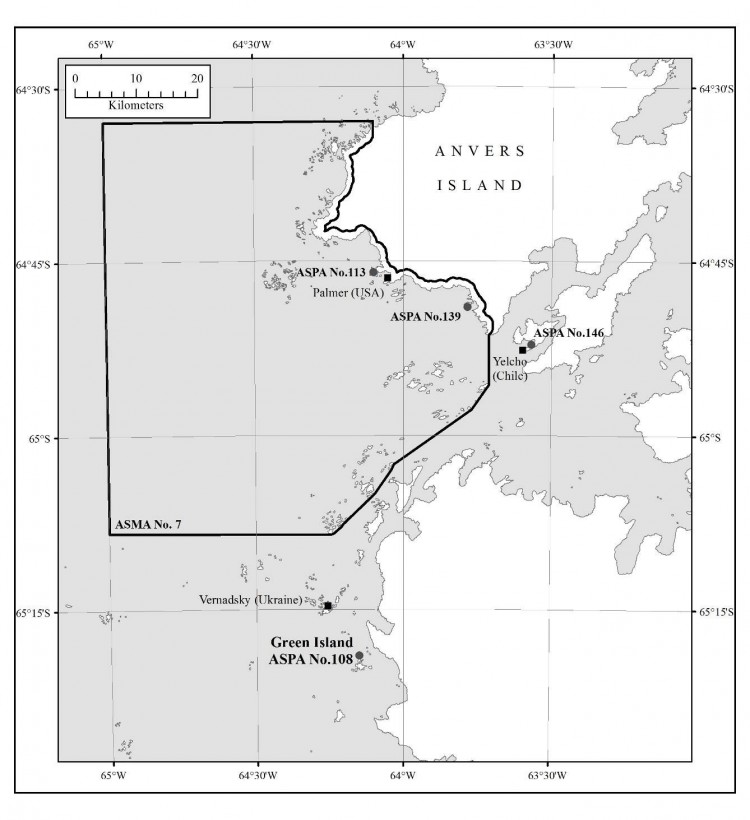
Carte 3. - ZSPA n° 108, île Green, îles Berthelot, péninsule antarctique, carte topographique. Carte réalisée à partir d'une étude de terrain le 24 février 2001 et d'une orthophotographie numérique (photographie aérienne de la British Antarctic Survey, prise le 14 février 2001). Spécifications de la carte - Projection : zone UTM 20S ; sphéroïde : WGS84 ; Datum : niveau moyen de la mer (EGM96).
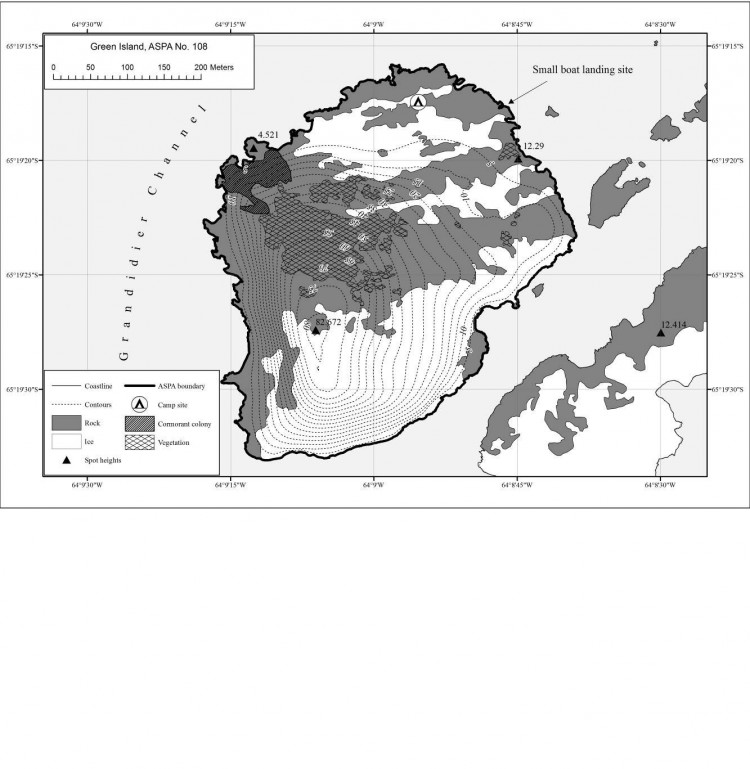
Annexe
MESURE 2 (2018)
ZONE SPÉCIALEMENT PROTÉGÉE DE L'ANTARCTIQUE N° 117 (ILE AVIAN, BAIE MARGUERITE, PÉNINSULE ANTARCTIQUE) : PLAN DE GESTION RÉVISÉ (ENSEMBLE UNE ANNEXE)
Les Représentants,
Rappelant les articles 3, 5 et 6 de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement, qui disposent de la désignation des zones spécialement protégées de l'Antarctique (« ZSPA ») et de l'adoption des plans de gestion pour ces zones ;
Rappelant :
- la recommandation XV-6 (1989), qui a désigné l'île Avian, nord-ouest de la baie Marguerite comme site présentant un intérêt scientifique particulier (« SISP ») n° 30 et a mis en annexe le plan de gestion du site ;
- la recommandation XVI-4 (1991), qui a rebaptisé le SISP n° 30 comme zone spécialement protégée (« ZSP ») n° 21 et a annexé un plan de gestion révisé pour la zone ;
- la décision 1 (2002) qui a renommé et renuméroté la ZSP n° 21 en ZSPA nº 117 ;
- la mesure 1 (2002) et la mesure 2 (2013), qui ont adopté des plans de gestion révisés pour la ZSPA n° 117 ;
Rappelant que les recommandations XV-6 (1989) et XVI-4 (1991) ne sont pas entrées en vigueur et ont été déclarées caduques par la Décision 1 (2011) ;
Notant que le Comité pour la protection de l'environnement a approuvé un plan de gestion révisé pour la ZSPA n° 117 ;
Souhaitant remplacer le plan de gestion actuel pour la ZSPA n° 117 avec le plan de gestion révisé ;
Recommandent à leurs gouvernements d'approuver la mesure suivante conformément au paragraphe 1 de l'article 6 de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement :
Que :
1. Le plan de gestion révisé pour la zone spécialement protégée de l'Antarctique n° 117 (île Avian, baie Marguerite, péninsule antarctique), qui figure en annexe de cette mesure, soit approuvé ; et que
2. Le plan de gestion pour la zone spécialement protégée de l'Antarctique n° 117 qui figure en annexe de la mesure 2 (2013) soit abrogé.
ANNEXE
PLAN DE GESTION POUR LA ZONE SPÉCIALEMENT PROTEGÉE DE L'ANTARCTIQUE N° 117
ÎLE AVIAN, BAIE MARGUERITE, PÉNINSULE ANTARCTIQUE
Introduction
La raison principale de la désignation de l'île Avian, baie Marguerite, péninsule antarctique (Lat. 67°46'S, Long. 68°54'O ; 0,49 km2) comme zone spécialement protégée de l'Antarctique (ZSPA) est la protection des valeurs environnementales de la zone et en particulier l'abondance et la diversité des oiseaux marins reproducteurs présents sur l'île.
L'île Avian est située au nord-ouest de la baie Marguerite, à 400 mètres au sud de l'île Adelaïde du côté est de la péninsule antarctique centrale. A l'origine, elle avait été désignée Site présentant un intérêt scientifique particulier (SISP) n° 30 dans la recommandation XV-6 (1989) sur proposition du Royaume-Uni. La zone comprenait l'île avec son littoral, mais excluait une petite zone près d'un refuge sur la côte nord-ouest de l'île. Les valeurs à protéger selon la désignation originale étaient les suivantes : l'abondance et la diversité des oiseaux marins reproducteurs présents sur l'île ; la colonie de pétrels géants (Macronectes giganteus) étant l'une des populations reproductrices connues de cette espèce situées le plus au sud ; et les cormorans impériaux (Phalacrocorax [atriceps] bransfieldensis) se reproduisant près de la limite méridionale de leur répartition. Par conséquent, il a été estimé que la zone était d'une valeur ornithologique exceptionnelle et qu'elle méritait une protection vis-à-vis des perturbations anthropiques inutiles.
La première désignation SISP de l'île Avian a été remplacée par une nouvelle désignation de zone spécialement protégée (ZSP), via la recommandation XVI-4 (1991, ZSP N° 21) sur proposition du Royaume-Uni.
Les limites de la ZSP étaient similaires à celles du SISP, mais incluaient la totalité de l'île et de la zone littorale, y compris le secteur près du refuge sur la côte nord-ouest de l'île. Après une nouvelle désignation, en tant que ZSPA n° 117 via la Décision 1 (2002), le Plan de gestion de la ZSPA a été approuvé dans la Mesure 1 (2002).
La zone s'inscrit dans le cadre plus large du système des zones protégées de l'Antarctique, en protégeant le site de reproduction de sept espèces d'oiseaux marins, y compris les pétrels géants, qui sont vulnérables aux perturbations. Aucune autre ZSPA dans la région ne protège une telle diversité d'espèces d'oiseaux en phase de reproduction. La résolution 3 (2008) recommandait que l'analyse des domaines environnementaux pour le continent antarctique serve de modèle dynamique pour l'identification des zones spécialement protégées de l'Antarctique dans le cadre environnemental et géographique systématisé visé à l'article 3 (2) de l'annexe V du Protocole (voir également Morgan et al., 2007). Selon ce modèle, l'île Avian relève essentiellement du domaine environnemental E (péninsule antarctique et principaux champs de glaces de l'île Alexander), que l'on trouve aussi dans les ZSPA 113, 114, 126, 128, 129, 133, 134, 139, 147, 149, 152 et dans les ZGSA 1 et 4. Néanmoins, l'île Avian étant majoritairement libre de glace, ce domaine ne représente peut-être pas pleinement le type d'environnement présent dans la zone. Bien qu'elle ne soit pas décrite en tant que telle dans Morgan et al., il est possible que l'île Avian soit mieux représentée par le domaine environnemental B (géologique des latitudes septentrionales moyennes de la péninsule antarctique). Parmi les autres zones protégées contenant le domaine environnemental B, on compte notamment les ZSPA 108, 115, 129, 134, 140 et 153 et la ZGSA 4. La ZSPA se trouve dans la Région de conservation biogéographique (RCBA) 3 : nord- ouest de la péninsule antarctique (Terauds et al., 2012 ; Terauds & Lee, 2016). Dans la résolution 5 (2015), les Parties ont reconnu l'intérêt des zones importantes pour la conservation des oiseaux de l'Antarctique (ZICO) dans la planification et le déploiement d'activités en Antarctique. La ZICO ANT095 île Avian a les mêmes frontières que la ZSPA n° 117 et a été désignée pour la protection du manchot Adélie (Pygoscelis adeliae), du cormoran impérial (Phalacrocorax [atriceps] bransfieldensis), et du labbe de McCormick (Stercorarius maccormicki).
1. Description des valeurs à protéger
Les valeurs environnementales exceptionnelles de la zone, qui constituent la raison principale de sa désignation comme ZSPA, sont les suivantes :
- la colonie de manchots d'Adélie (Pygoscelis adeliae) est l'une des plus importantes sur la terre de Palmer, comptant 77 515 couples reproducteurs ;
- la colonie de cormorans impériaux (Phalacrocorax [atriceps] bransfieldensis) est l'un des sites de reproduction connus les plus vastes en Antarctique et elle est proche de la limite méridionale de la répartition de cette espèce ;
- le caractère unique de l'île : elle est le seul site connu dans la péninsule antarctique où sept espèces d'oiseaux marins se reproduisent avec une telle proximité les unes des autres dans l'espace restreint d'une seule île de petite taille et à la densité de population exceptionnellement élevée, l'île entière étant occupée par des oiseaux reproducteurs pendant l'été ;
- la colonie de pétrels géants (Macronectes giganteus) est l'une des deux plus importantes de la péninsule antarctique ;
- la colonie de goélands dominicains (Larus dominicanus) est également importante et ils se reproduisent près de la limite sud de leur répartition ; et
- l'espèce de mousse Warnstorfia fontinaliopsis sur l'île d'Avian est proche de la frontière méridionale de sa répartition connue.
2. Buts et objectifs
Les objectifs du présent plan de gestion sont les suivants :
- éviter toute détérioration ou tout risque de détérioration des valeurs de la zone en empêchant toute perturbation humaine inutile ;
- éviter ou réduire au maximum l'introduction de plantes, d'animaux et de microbes non indigènes dans la zone ;
- limiter le risque d'introduction d'agents pathogènes qui pourraient provoquer des maladies au sein des populations fauniques de la zone ;
- permettre d'effectuer des recherches scientifiques dans la zone, pour autant qu'elles soient indispensables, qu'elles ne puissent être menées ailleurs et qu'elles ne portent pas atteinte à l'écosystème naturel de la zone ; et
- préserver l'écosystème naturel de la zone afin que celle-ci serve de zone de référence pour les études futures.
3. Activités de gestion
Les activités de gestion ci-dessous seront menées à bien afin de protéger les valeurs de la zone :
Un exemplaire de ce plan de gestion sera mis à disposition dans la station Teniente Luis Carvajal (Chili ; Lat. 67°46'S, Long. 68°55'O), la base antarctique Rothera (R.-U. ; Lat. 67°34'S, Long. 68°07'O) et la station General San Martín (Argentine ; Lat. 68°08'S, Long. 67°06'O).
Le plan de gestion sera réexaminé au moins tous les 5 ans et mis à jour en conséquence.
Les équipes de terrain en visite seront pleinement informées par l'autorité nationale sur les valeurs devant être préservées dans la zone et sur les mesures de précaution et d'atténuation décrites dans le présent plan de gestion.
Toutes les activités scientifiques et de gestion entreprises au sein de la zone devraient faire l'objet d'une évaluation d'impact sur l'environnement conformément à ce que requiert l'annexe I du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement.
Des copies du présent plan de gestion doivent être mises à la disposition des navires prévoyant de visiter les abords de la zone.
Tous les pilotes qui opèrent dans la région seront tenus informés de l'emplacement, des limites et des restrictions applicables à l'entrée et au survol de la zone.
Les bornes, panneaux et autres structures érigés à l'intérieur de la zone à des fins scientifiques ou de gestion seront correctement fixés, maintenus en bon état et enlevés lorsqu'ils ne seront plus nécessaires.
Conformément aux dispositions de l'annexe III du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement, les équipements ou matériels abandonnés seront enlevés dans toute la mesure du possible, à condition que leur enlèvement n'ait pas d'impact préjudiciable sur l'environnement et les valeurs de la zone.
Les directeurs des programmes antarctiques nationaux en cours d'exécution dans la région se livreront entre eux à des consultations pour veiller à ce que les activités de gestion susmentionnées soient mises en œuvre.
4. Durée de désignation
La zone est désignée pour une période indéterminée.
5. Cartes et photographies
Carte 1. - Ile Avian, ZSPA n° 117, par rapport à la baie Marguerite, indiquant l'emplacement des stations Teniente Luis Carvajal (Chili), Rothera (R.-U.) et General San Martín (Argentine). Les emplacements d'autres zones protégées comprises dans la baie Marguerite (ZSPA n° 107 sur l'île Emperor (îles Dion), ZSPA n° 115 sur l'île Lagotellerie et ZSPA n° 129 sur la pointe Rothera) sont également indiqués. Encart : emplacement de l'île Avian sur la péninsule antarctique.
Carte 2. - Ile Avian (ZSPA n° 117), carte topographique. Spécifications de la carte - projection : conique conforme de Lambert ; parallèles types : 1er 67°30'00”S ; 2e 68°00'00”S ; méridien central : 68°55'00”O ; latitude d'origine : 68°00'00”S ; sphéroïde : WGS84 ; datum : niveau moyen de la mer ; équidistance des courbes de niveau verticales : 5 m ; précision horizontale : ± 5 m ; précision verticale ± 1,5 m.
Carte 3. - Ile Avian (ZSPA n° 117), croquis cartographique de la faune en phase de reproduction. La position des nids et des colonies sont d'une précision de ± 25 m. Ces informations proviennent de Poncet (1982). Spécifications de la carte - projection : conique conforme de Lambert ; parallèles types : 1er 67° 30' 00”S ; 2e 68°00'00”S ; méridien central : 68°55'00”O ; latitude d'origine : 68°00'00"S ; sphéroïde : WGS84 ; datum : niveau moyen de la mer ; équidistance des courbes de niveau verticales : 5 m ; précision horizontale : ± 5 m ; précision verticale ± 1,5 m.
6. Description de la zone
6 (i) Coordonnées géographiques, bornage et milieu naturel
Description générale
L'île Avian (Lat. 67°46'S ; Long. 68°54'O, 0,49 km2) est située au nord-ouest de la baie Marguerite, à 400 mètres au sud de l'extrémité sud-ouest de l'île Adélaïde (Carte 1). L'île fait environ 1,45 km de long sur 0,8 km en son point le plus large et s'élève à environ 65 mètres d'altitude. Sa forme est plutôt triangulaire. Elle est rocheuse avec un relief assez bas qui ne dépasse pas les 10 mètres au nord mais qui atteint 30 mètres au centre et 40 mètres au sud où plusieurs versants rocheux et glacés pouvant atteindre 30 mètres rejoignent la mer. La côte est irrégulière et rocheuse avec de nombreux îlots proches mais elle comporte également plusieurs plages accessibles sur les côtes nord et est. L'île est en général libre de glace en été. Son habitat est particulièrement adapté à une multitude d'espèces d'oiseaux reproducteurs : les pentes exposées au nord et bien drainées conviennent parfaitement aux cormorans impériaux (Phalacrocorax [atriceps] bransfieldensis) ; les rochers fragmentées et les blocs rocheux fissurés sont propices aux petits oiseaux nicheurs tels que les océanites de Wilson (Oceanites oceanicus) ; les hauteurs rocailleuses plus élevées sont recherchées par les pétrels géants (Macronectes giganteus) ; et les plus vastes étendues libres de glace sont convoitées par les manchots d'Adélie (Pygoscelis adeliae). La présence de ces derniers attire les labbes (Stercorarius maccormicki et Stercorarius antarcticus) et les goélands dominicains (Larus dominicanus).
Limites
La zone désignée comprend la totalité de l'île Avian, sa zone littorale, des îlots et des rochers alentours ainsi qu'une zone tampon abritant le milieu marin avoisinant (y compris les glaces marines lorsqu'il y en a) sur une distance de 100 mètres autour du rivage de l'île principale (Carte 2). Aucun dispositif de bornage n'a été installé car le littoral à lui seul détermine clairement la limite marine.
Climat et glace marine
Il n'existe aucune donnée d'archive détaillée sur la météorologie pour l'île Avian mais les relevés effectués entre 1962 et 1974 pour la base Adélaïde (appartenant autrefois au Royaume-Uni et aujourd'hui au Chili qui l'a renommée station Teniente Luis Carvajal), distante de 1,2 km, indiquent une température journalière maximum moyenne de 3 °C en février (record maximum de 9 °C) et une température journalière minimum moyenne de - 8 °C en août (record minimum de - 44 °C). Le même cycle global a été constaté lors d'observations effectuées sur l'île tout au long de l'année en 1978-1979 (Poncet et Poncet, 1979). Cette année-là, les précipitations sur l'île se sont présentées sous forme de neige et sont surtout tombées du mois d'août au mois d'octobre, même si des pluies et des chutes de neige sporadiques ont été enregistrées durant l'été.
La baie Marguerite peut geler en hiver bien que l'étendue et les caractéristiques de la glace marine montrent des variations considérables d'une saison à l'autre. Malgré l'étendue et la persistance fréquente de la glace marine dans la région, une polynie se forme de manière récurrente à proximité de l'île Avian, ce qui peut permettre l'absence de glace dans cette zone à partir d'octobre. En outre, de forts courants de marée autour de l'île permettent de maintenir les eaux environnantes libres de glace durant la majeure partie de l'année, ce qui facilite l'accès aux aires d'alimentation pour plusieurs espèces. L'île n'est pas spécialement balayée par les vents dont la force est évaluée en moyenne annuelle à 10 nœuds (1978-1979). Cependant, les vents catabatiques violents qui descendent de l'île Adélaïde et soufflent de un à trois jours quelques fois par mois, réduisent l'accumulation de la neige sur l'île et éloignent la glace marine du littoral, contribuant ainsi à la formation de polynies. Cet enneigement relativement faible joue un rôle capital dans l'établissement de colonies par les oiseaux.
Géologie, géomorphologie et sols
La roche mère de l'île Avian fait partie d'un bloc faillé vers le bas à l'extrémité sud-ouest de l'île Adélaïde. Elle est constituée de grès volcaniclastiques interstratifiés riches en feldspath et en lithique. La zone contient également du grès tuffacé stratifié, du grès graveleux riche en lithique volcanique et des brèches granuleuses volcaniques. Ces dernières sont probablement un dépôt volcanique primaire tandis que les autres éléments de la séquence sont en grande partie composés de matières volcaniques reformées. Ladite séquence fait partie de la formation du mont Liotard, situé dans l'île Adélaïde, et date probablement de la période du Crétacé tardif (Griffiths, 1992 ; Moyes et al, 1994 ; Riley et al., 2012). A l'exception des affleurements rocheux, la surface est principalement constituée de roches brisées par la glace et de pergélisol. Les sols ornithogéniques sont très répandus, surtout au nord, mais les sols constitués de tourbe organique sont pratiquement absents et, lorsqu'ils existent, ils sont peu développés et associés à la croissance de la mousse. Plusieurs plages surélevées ont été répertoriées sur l'île Avian, mais le reste de la géomorphologie n'a fait l'objet d'aucune description.
Cours d'eau et lacs
L'île Avian compte plusieurs étangs d'eau douce éphémères allant jusqu'à 10 000 m2 et 40 cm de profondeur, les deux plus grands étant situés sur la côte est, à environ 5 mètres d'altitude, et sur la côte nord-ouest près du niveau de la mer. De nombreux petits bassins et canaux d'eau de fonte font leur apparition lors de la fonte des neiges saisonnières et de petits cours d'eau canalisent l'eau des vallées à proximité des étangs. Tant les étangs que les bassins d'eau de fonte gèlent en hiver. Les guanos, source d'éléments nutritifs, enrichissent organiquement les étendues d'eau douce sur l'île, et en été de nombreux étangs abritent une flore et une faune benthiques très riches d'algues, de phyllopodes, de copépodes, de nématodes, de protozoaires, de rotifères et de tardigrades. Un grand nombre de crustacés de l'espèce Branchinecta a également été observé (Poncet et Poncet, 1979). L'écologie des eaux douces de l'île n'a pas fait l'objet d'une étude détaillée.
Oiseaux nicheurs
Sept espèces d'oiseaux se reproduisent sur l'île Avian. Ce chiffre est relativement élevé comparé à celui d'autres sites de la péninsule antarctique. Plusieurs espèces ont des populations particulièrement élevées à tel point que certaines d'entre elles sont les plus nombreuses de la région de la péninsule antarctique (Carte 3). Des informations détaillées pour toutes les espèces ont été recueillies tout au long de l'année en 1978-1979 (Poncet et Poncet, 1979) mais, à part ces données, les informations restent sporadiques. Par conséquent, les descriptions ci-dessous sont axées sur des observations réalisées au cours d'une seule saison et il convient de souligner que ces données ne sont pas nécessairement représentatives de l'évolution des tendances démographiques à long terme. Toutefois, il s'agit des données les plus fiables disponibles à l'heure actuelle.
La colonie de manchots Adélie (Pygoscelis adeliae) occupe la moitié nord et le centre de la côte orientale de l'île (Carte 3). Le premier plan de gestion indiquait que cette colonie était « la plus importante de la péninsule antarctique puisqu'elle représentait un tiers de la population totale en phase de reproduction dans la région ». Bien qu'aucune donnée récente ne permette de corroborer cette observation (une colonie de la péninsule antarctique se compose de plus de 120 000 couples [Woehler, 1993] ), la colonie de l'île Avian constitue une des plus importantes populations en phase de reproduction de la terre de Palmer. Des recherches récentes tendent à montrer que le nombre de manchots Adélie est en diminution dans presque tous les endroits de la péninsule antarctique (Lynch et al., 2012). Selon les dernières estimations de population pour les manchots Adélie, datant de la saison 2015-2016, l'île Avian abriterait 65 888 couples reproducteurs (W. Fraser, comm. pers. 2018). Deux ensembles de données disponibles sur les manchots Adélie présents sur l'île Avian collectées en 2013 indiquaient que la population de couples en phase de reproduction s'élevait à 77 515 en janvier 2013 (± 5 %) (W. Fraser, comm. pers. 2013 ; Sailley et al., 2013), et le nombre de couples à 47 146 (Casanovas et al., 2015), bien que l'écart entre ces nombres ne soit pas clairement expliqué. Ces données sont comparables avec une estimation du nombre de manchots Adélie, fondée sur des photographies aériennes prises en décembre 1998, qui ont révélé 87 850 oiseaux (± 0.16 S.D. ; Woehler, 1993), tandis qu'en date du 11 novembre 1978, leur population s'élevait à 36 500 (Poncet et Poncet, 1979).
En 1978-1979, les manchots Adélie ont été vus sur l'île d'octobre à la fin du mois d'avril, la ponte des œufs ayant lieu d'octobre à novembre et les premières éclosions aux environs de la mi-décembre. L'activité des bébés manchots a été observée dès la mi-janvier et les premiers oisillons sont devenus indépendants dès la fin de ce même mois. La plupart des adultes en mue et des bébés manchots indépendants ont quitté l'île dès la troisième semaine de février, même si certains groupes sont revenus périodiquement aux mois de mars et d'avril.
Une grande colonie de cormorans impériaux (Phalacrocorax [atriceps] bransfieldensis) a été observée dans trois groupes situés sur la côte sud-ouest à l'extrémité de l'île (Carte 3). Cependant, lors d'une visite effectuée les 26 et 27 janvier 2011, il est apparu que les deux sites de colonies situés le plus au nord n'étaient pas occupés et que les buttes de nidification étaient en mauvais état. Ces observations laissent à penser que ces sites étaient probablement abandonnés depuis un certain temps. Stonehouse (1949) avait rapporté la présence d'environ 300 oiseaux en octobre 1948 et un nombre similaire d'individus avait été enregistré à la mi-novembre 1968, dont la plupart était en phase de reproduction (Willey, 1969). Poncet et Poncet (1979) ont observé 320 couples en 1978 et environ 670 couples le 17 janvier 1989 (Poncet, 1990). Un recensement effectué le 23 février 2001 faisait état de 185 jeunes cormorans, mais il est probable que certains d'entre eux avaient déjà quitté l'endroit au moment du recensement. Quelque 250 sites de nidification ont été répertoriés. Le recensement réalisé de la mi-janvier à la fin janvier 2013 faisait état de 302 couples en phase de reproduction (W. Fraser, comm. pers., 2013). En 1968, des cormorans impériaux ont été observés sur l'île à partir du 12 août, la ponte intervenant à partir du mois de novembre et l'éclosion au mois de décembre (Willey, 1969). En 1978-1979, ils ont été observés de septembre à juin, la ponte intervenant de novembre à janvier, jusqu'aux premières éclosions, et les tout jeunes cormorans ont commencé à devenir indépendants pendant la troisième semaine de février (Poncet et Poncet, 1979).
Parmi les colonies de pétrels géants (Macronectes giganteus) connues au sud des îles Shetland du Sud, l'île Avian est l'une des deux plus importantes. Elle abrite une partie importante de la population en phase de reproduction de la région de la péninsule antarctique méridionale (avec une estimation de 1190 couples en 1999-2000 ; Patterson et al., 2008). En 1979, les pétrels géants occupaient principalement les affleurements rocheux élevés des parties méridionale et centrale de l'île, se répartissant en quatre groupes principaux (Carte 3). Des données sur le nombre d'oiseaux présents sur l'île figurent dans le Tableau 1.
Tableau 1. - Nombre de pétrels géants (Macronectes giganteus) sur l'île Avian
|
Année |
Nombre d'oiseaux |
Nombre de couples |
Nombre d'oisillons |
Source |
|
1948 |
~ 100 |
nd |
nd |
Stonehouse, 1949 |
|
1968 |
400 |
163 |
nd |
Willey, 1969 |
|
1979 |
nd |
197 |
nd |
Poncet & Poncet, 1979 |
|
1989 |
nd |
250 |
nd |
Poncet, 1990 |
|
2001 |
nd |
nd |
237 |
Harris, 2001 |
|
2013 |
nd |
470 |
nd |
W. Fraser, comm. pers., 2013 |
|
nd - non disponible. |
En 1978-1979, les oiseaux étaient présents sur l'île Avian entre la mi-septembre et le mois de juin. Pendant cette saison, la ponte s'étendait de la fin du mois d'octobre à la fin du mois de novembre, l'éclosion intervenant pendant le mois de janvier et les jeunes oiseaux acquérant généralement leur indépendance au plus tard en avril. Durant l'été austral de 1978-1979, jusqu'à 100 pétrels non reproducteurs ont été observés sur l'île lors de la pariade en octobre mais ce nombre a été réduit à quelques oiseaux au fil de la saison.
Environ 200 goélands dominicains (Larus dominicanus) adultes, dont 60 couples en phase de reproduction, ont été observés sur l'île Avian en 1978-1979. Ces oiseaux étaient répartis sur toute l'île mais se retrouvaient principalement au centre et au sud sur le relief (Poncet et Poncet, 1979) (Carte 3). Lors de l'été austral 1978- 1979, la majorité des goélands en phase de reproduction sont arrivés au début du mois d'octobre, la ponte intervenant à la mi-novembre et l'éclosion un mois plus tard. Aucune donnée détaillée n'est disponible compte tenu des risques de perturbation anthropique inhérents aux opérations de collecte de données, qui affecteraient fortement la reproduction de ces espèces. Cependant, à peine 12 jeunes goélands ont été observés sur l'île à la fin du mois de janvier 1979, ce qui indique que le taux de reproduction a été faible lors de cette saison. La cause exacte de cet état de fait - perturbation humaine ou facteurs naturels - n'a pas pu être déterminée. En 1967, 19 couples et entre 80 et 120 oiseaux ont été répertoriés (Barlow, 1968).
Une estimation effectuée en 1978-1979 confirme la présence sur l'île d'au moins quelques centaines de couples d'océanites de Wilson (Oceanites oceanicus) en phase de reproduction (Poncet et Poncet, 1979). Ils ont été observés dès la deuxième semaine de novembre, la ponte et l'incubation intervenant probablement jusqu'à la mi-décembre. A la fin du mois de mars, la majeure partie des adultes et des jeunes déjà indépendants avait quitté l'endroit. La plupart des affleurements rocheux de la moitié nord de l'île, ainsi que toutes les pentes rocheuses stables du sud, constituent un habitat idéal pour cette espèce.
En 1978-1979, environ 25 à 30 couples de labbes de McCormick (Stercorarius maccormicki) étaient en phase de reproduction sur l'île Avian. Leurs nids étaient largement dispersés sur l'île mais la plupart se trouvaient au centre et à l'est, notamment sur les pentes surplombant la colonie de manchots Adélie (Carte 3). D'importants groupes d'oiseaux non reproducteurs (environ 150 ; Poncet et Poncet, 1979) ont été observés autour d'un lac peu profond sur le flanc est de l'île. En 1968, Barlow a observé quelque 200 labbes qui n'étaient pas en phase de reproduction. En 2004, 195 couples de labbes de McCormick en phase de reproduction ont été enregistrés dans les parties centrale et orientale de l'île (W. Fraser, comm. pers., 2015), en plus des 880 labbes non reproducteurs également observés sur l'île (W. Fraser pers. comm. 2015, dans une rectification des données renseignées dans Ritz et al., 2006). Lors de l'été austral de 1978-1979, ils se sont installés sur l'île à la fin du mois d'octobre, la ponte intervenant au début de décembre et l'éclosion se terminant à la fin du mois de janvier. Les adultes et les jeunes devenus indépendants avaient en général quitté les lieux à la fin du mois de mars, certains reproducteurs tardifs restant sur les lieux jusqu'à la mi-avril. Au cours de cet été austral, la phase de reproduction a donné le résultat d'un jeune labbe par nid. Barlow (1968) a observé 12 couples de labbes antarctiques (Stercorarius antarcticus), bien que ce nombre puisse inclure des labbes de McCormick. Un couple de labbes antarctiques a été observé au sud-ouest de l'île au cours de l'été austral 1978-1979. Cette espèce n'avait jamais été observée à une latitude aussi méridionale de la péninsule antarctique. La présence de plusieurs labbes antarctiques non reproducteurs a également été signalée pendant la même saison.
Plusieurs autres espèces d'oiseaux qui se reproduisent ailleurs dans la baie Marguerite visitent fréquemment l'île Avian. Il s'agit notamment de la sterne couronnée (Sterna vittata), du pétrel des neiges (Pagodroma nivea) et du fulmar austral (Fulmarus glacialoides). Aucune nidification de ces espèces n'a été observée sur l'île. Quelques pétrels antarctiques (Thalassoica antarctica) ont été vus occasionnellement. La présence du damier du cap (Daption capense) a été signalée en octobre 1948 (Stonehouse, 1949). Des manchots royaux (Aptenodytes patagonicus) et à jugulaire (Pygoscelis antarctica) solitaires ont été observés en 1975 et en 1989, respectivement.
Biologie terrestre
La végétation de l'île Avian est en général clairsemée et la flore n'a pas fait l'objet d'une étude détaillée. Il n'existe pas de phanérogames sur l'île et les cryptogames sont en quantité limitée même si la flore de lichens est abondante. A ce jour, 9 espèces de mousses et 11 de lichens ont été identifiées dans la zone.
Ces mousses sont les suivantes : Andreaea depressinervis, Brachythecium austrosalebrosum, Bryum argenteum, B. pseudotriquetrum, Ceratodon purpureus, Pohlia cruda, P. nutans, Sanionia georgico- uncinata, S. uncinata, Syntrichia magellanica et Warnstorfia fontinaliopsis. Cette dernière espèce se retrouve à la limite sud de sa zone de répartition connue sur l'île Avian (Smith, 1996). Les mousses se développent uniquement aux endroits de l'île qui ne sont occupés ni par les manchots Adélie ni par les cormorans impériaux, dans les zones humides ou à proximité des bassins d'eau de fonte. Des concentrations éparses de mousses allant jusqu'à 100 m2 entourent les berges d'un petit étang sur la colline située au sud de la zone, à environ 30 mètres d'altitude. L'algue verte Prasiola crispa est très répandue dans les zones humides de l'île et une espèce d'hépatique, Cephaloziella varians, a également été identifiée.
Les lichens identifiés sur l'île sont les suivants : Acarospora macrocyclos, Cladonia fimbriata, C. gracilis, Dermatocarpon antarcticum, Lecanora dancoensis, Lecidea brabantica, Physcia caesia, Rinodina egentissima, Siphulina orphnina, Thamnolecania brialmontii et Usnea antarctica. Les communautés les plus importantes se trouvent sur les affleurements rocheux au sud de l'île.
La faune de microinvertébrés, les champignons et les bactéries de l'île Avian doivent encore faire l'objet d'une étude approfondie. A ce jour, seul l'acarien mésostigmatide (Gamasellus racovitzai) a été décrit (base de données sur les invertébrés de la British Antarctic Survey, 1999), bien qu'un collembole et plusieurs autres espèces d'acariens aient été observés sans toutefois être identifiés (Poncet, 1990). Quelques espèces de nématodes (dominées par l'espèce Plectus) (Spaull, 1973) et un champignon (Thyronectria hyperantarctica) (base de données sur les invertébrés de la British Antarctic Survey, 1999) ont été enregistrés sur l'île.
Mammifères en phase de reproduction et écosystème marin
En 1978-1979, des phoques de Weddell (Leptonychotes weddellii) étaient souvent présents sur l'île Avian et ses alentours. Pendant l'hiver, plus d'une douzaine d'entre eux sont restés sur place et se sont installés sur la glace côtière (Poncet, 1990). Plusieurs bébés phoques sont nés sur le littoral de l'île pendant la dernière semaine de septembre 1978. Un éléphant de mer (Mirounga leonina) a été observé s'occupant de sa progéniture sur la côte nord-est de l'île Avian le 10 octobre 1969 (Bramwell, 1969). Des photos aériennes prises le 15 décembre 1998 ont montré 182 éléphants de mer regroupés principalement à proximité des étangs. Des léopards de mer (Hydrurga leptonyx) ont été observés sur le littoral, dont un sur la côte, durant l'hiver de 1978. Un certain nombre d'otaries de Kerguelen (Arctocephalus gazella), qui n'étaient pas en phase de reproduction, a été observé sur l'île en mars 1997 (Gray et Fox, 1997), à la fin de janvier 1999 (Fox, comm. pers., 1999) et en janvier 2011. Au moins plusieurs centaines de ces otaries étaient présentes lors des observations du 23 février 2001 (Harris, 2001), en particulier sur les plages et les endroits de faible altitude du centre et du nord de l'île. Des phoques crabiers (Lobodon carcinophagus) sont régulièrement observés dans la baie Marguerite mais n'ont pas été aperçus sur l'île Avian. L'écosystème marin autour de l'île Avian n'a fait l'objet d'aucune étude.
Activités humaines/impacts
Les activités humaines sur l'île Avian ont été sporadiques. Les archives indiquent que la première visite a eu lieu en octobre 1948 lorsque les membres de l'expédition britannique sur l'île Stonington ont découvert la grande colonie de manchots Adélie sur l'île Avian (qui à l'époque était une des îles Henkes). Les visites ultérieures ont été organisées à des fins scientifiques, récréatives (personnel de station), touristiques et logistiques (recensements, etc.). Des refuges ont été installés sur l'île respectivement en 1957 et en 1962 par l'Argentine et le Chili. [cf. section 6 (iii)].
En novembre 1968, une expédition géologique de deux personnes a campé pendant une dizaine de jours au sud-est de l'île (Elliott, 1969). Cette même année, une expédition navale d'exploration hydrographique du Royaume-Uni a établi son campement sur la côte est de l'île durant l'été. Les amarres et les chaînes permanentes pour le mouillage des vaisseaux de l'expédition ont été installées dans une petite baie sur la côte nord-ouest et ces dispositifs étaient encore là en 1989 (Poncet, 1990).
En 1969, une autre expédition a établi pendant un mois son campement sur l'île pour mener des recherches sur le virus du rhume : des chiens faisant partie de l'expérience ont été inoculés et ramenés à la base (Bramwell, 1969). Ces animaux accompagnaient souvent le personnel lors des visites régulières sur l'île Avian pendant la période d'activités menées par le personnel de la base britannique installée sur l'île Adélaïde, mais les impacts liés à ces activités sont inconnus.
En 1978-1979, un groupe de deux personnes a passé un an sur l'île, à bord du bateau de plaisance Damien II, afin de procéder à des observations détaillées de l'avifaune et d'étudier d'autres aspects de la biologie et de l'écosystème de l'île (Poncet et Poncet, 1979 ; Poncet, 1982 ; Poncet, 1990). Le bateau était amarré dans une petite anse sur la côte nord-ouest. Les chercheurs ont régulièrement visité l'île au cours des dix années suivantes avant qu'elle ne soit désignée zone spécialement protégée.
Des activités de cartographie et de photographie aérienne ont été réalisées sur l'île ainsi que dans son espace aérien en 1996-1998 (Fox et Gray, 1997 ; Gray et Fox, 1997) et en 1998-1999 (Fox, comm. pers., 1999).
Les impacts de ces activités n'ont pas été décrits et restent inconnus mais ils semblent avoir été relativement mineurs et s'être limités à la perturbation passagère des oiseaux en phase de reproduction, à l'installation des campements, aux traces de pas, à l'abandon occasionnel de détritus, aux déchets humains, aux prélèvements d'échantillons à des fins scientifiques et à l'installation de balises. Malgré la nature transitoire de la majorité de ces facteurs de perturbation, il a été rapporté que des visites humaines sur le site avaient entraîné la destruction d'œufs et la mort d'oisillons, soit par abandon du nid, soit par prédation opportuniste.
Plusieurs espèces telles que le pétrel géant et le goéland dominicain sont particulièrement vulnérables à toute perturbation et des observations ont révélé qu'ils abandonnaient leur nid à des périodes spécifiques du cycle de nidification s'ils voyaient l'homme s'approcher à moins de 100 mètres (Poncet, 1990). Quelque 140 personnes, y compris une embarcation touristique de 100 personnes, ont été observées en visite sur l'île Avian au cours de l'été 1989-1990. Le nombre et le caractère non réglementée de ces visites ont suscité une inquiétude croissante et ont contribué à la désignation de l'île Avian comme ZSP.
Les impacts les plus visibles et durables sont liés à la présence des deux refuges et des deux phares décrits à la section 6 (iii) qui sont situés à proximité d'oiseaux en phase de reproduction. Les deux refuges étaient toujours en mauvais état en février 2001 et lors d'une visite de gestion environnementale en janvier 2011 et janvier 2016 où de nouvelles détériorations ont été signalées. A ces mêmes dates et en février 2011, des oiseaux et des phoques ont été observés dans les ordures autour des refuges. Le refuge érigé sur la côte est (Lat. 67°46'26 ”S ; Long. 68°53'01”O) en 1957 a été exposé aux intempéries : la porte, ayant été arrachée de ses charnières, est tombée et la base du mur sud du refuge a désormais un grand trou (à peu près 0,25 m2). Des boîtes rouillées et du verre brisé ont été trouvés par terre. Des métaux rouillés (notamment des bardages en tôle ondulée, des jalons et des haubans), des fragments de bois en décomposition et du verre brisé ont également été trouvés à proximité du refuge. Un fût de carburant, d'une contenance de 205 litres, corrodé et vide, se trouve au sud du refuge.
En janvier 2011, le plus grand refuge, installé sur la côte nord-ouest (Lat. 67°46'08"S ; Long. 68°53'29"O) en 1962, était également en mauvais état. Il s'est sensiblement détérioré à cause de l'humidité ; ses poutres se sont déformées et des surfaces importantes de moisissure et d'algues s'étendent sur les murs et sur le revêtement du plafond. Une grande partie du plafond s'est effondrée laissant apparaitre le toit. En janvier 2016, il a été observé que des travaux avaient été entrepris pour éviter de nouvelles dégradations (les fenêtres et les portes ont par exemple été murées).
Le plus ancien des deux phares n'est plus en service et sa structure métallique, bien que toujours en place, affiche un état d'oxydation et de détérioration avancé. Le nouveau phare, mis en place en février 1998, semblait être en bon état en janvier 2011.
6 (ii) Accès à la zone
L'accès à la zone par petites embarcations est autorisé aux endroits prévus à cet effet sur la côte nord-ouest centrale (Lat. 67°46'08,1”S ; Long. 68°53'30,1”O) ou sur la côte est centrale de l'île (Lat. 67°46'25,5”S ; Long. 68°52'57,0”O) (Carte 2). Si les conditions en mer ou l'état de la glace compliquent l'accès, les petites embarcations peuvent accéder au site en d'autres points de la côte si la situation du moment le permet.
L'accès au site en véhicule est autorisé lorsque la côte est entourée de glace marine et, dans ce cas, tout véhicule doit utiliser ces points d'accès et être garé sur le littoral.
Les déplacements en véhicule ou en petite embarcation dans la partie marine de la zone ne sont pas soumis à un itinéraire particulier mais doivent suivre l'itinéraire le plus court en fonction des objectifs et des exigences des activités autorisées.
L'équipage ou les autres personnes dans les véhicules ou les embarcations ne peuvent en aucun cas se déplacer à pied dans les alentours immédiats du site de débarquement sauf avis contraire stipulé dans le permis.
Tout aéronef doit éviter d'atterrir dans la zone quel que soit le moment de l'année.
Un permis peut être délivré pour accéder à la zone en hélicoptère quand cela s'avère nécessaire pour répondre à des objectifs essentiels, tels que l'installation, l'entretien ou l'enlèvement de structures, et s'il n'existe aucun autre moyen. Dans ce cas-là, la nécessité de l'accès en hélicoptère, les alternatives et le degré de perturbation potentielle des oiseaux en phase de reproduction doivent être évalués avec précision avant la délivrance du permis. Ce permis définira clairement les conditions d'accès par hélicoptère sur la base des conclusions de l'évaluation.
6 (iii) Structures à l'intérieur et à proximité de la zone
Deux petits refuges désaffectés et deux phares sont présents dans la zone. Un refuge installé par le Chili en 1962 est situé sur la côte nord-ouest de l'île (Lat. 67°46'08”S ; Long. 68°53'29”O). L'autre refuge, construit par l'Argentine en 1957, est à 650 mètres au sud-ouest de cet emplacement, sur la côte est de la zone (Lat. 67°46'26”S ; Long. 68°53'01"O). Les deux refuges étaient en mauvais état en janvier 2016. Des réparations ont été effectuées afin de prévenir toute détérioration supplémentaire. Eu égard à la date de construction du refuge argentin, laquelle précède la signature du Traité sur l'Antarctique, l'Argentine va réévaluer la valeur historique potentielle de ce qu'il reste du refuge. Des mesures seront ensuite prises afin d'en conserver les valeurs historiques et de s'assurer que le refuge ne nuise pas à l'environnement.
Une vieille structure en fer forgé qui pourrait avoir été installée par le Royaume-Uni lors de l'opération de la base Adélaïde et avoir servi de guide à la navigation, est située à environ 38 mètres du sommet de l'île (Lat. 67°46'35,5”S ; Long. 68°53'25,2”O). Cette structure est toujours en place, bien qu'elle montre un état d'oxydation certain.
Un nouveau phare a été mis en place par le Chili en février 1998 sur un site adjacent, à la même altitude (Lat. 67°46'35,3”S ; Long. 68°53'26,0”O). Il s'agit d'une solide structure métallique de forme cylindrique, d'environ 2 mètres de diamètre et 2,50 mètres de haut, protégée par une couche de peinture et fixée sur un socle en béton d'environ 2,50 mètres sur 2,50 mètres. Un phare, des rails de protection et des panneaux solaires sont fixés au sommet de la structure. Il ne semble pas y avoir d'autres structures sur l'île.
Quatre bornes de contrôle ont été installées sur l'île le 31 janvier 1999 (Carte 2). La borne la plus au sud est un clou d'arpentage installé dans la roche mère et recouvert d'un cairn et est située à côté du phare. Une autre borne similaire est installée au point culminant de la crête inférieure de la côte nord-est de l'île et est également couverte d'un cairn. Les deux dernières bornes, également des clous d'arpentage, sont fixées au toit de chacun des refuges.
La station de recherche scientifique la plus proche se trouve à 1,2 km au nord-ouest. Il s'agit de la station Teniente Luis Carvajal (Chili) située dans la partie méridionale de l'île Adélaïde (Lat. 67°46'S ; Long. 68°55'O). Depuis 1982, cette installation fonctionne uniquement en été et elle est ouverte d'octobre à mars. Pendant cette période, elle accueille en général jusqu'à dix personnes. Elle a été au départ installée par le Royaume-Uni qui l'a administrée en permanence de 1961 à 1977.
6 (iv) Emplacement d'autres zones protégées à proximité
Les autres zones protégées proches de l'île Avian sont :
- la ZSPA n° 107, île Empereur, îles Dion, baie Marguerite, péninsule antarctique, (Lat. 67°52'S ; Long. 68°42'O), à 12,5 km au sud-sud-est ;
- la ZSPA n° 129, pointe Rothera, île Adélaïde, (Lat. 67°34'S ; Long. 68°08'O), à 40 km au nord-est ; et
- la ZSPA n° 115, île Lagotellerie, baie Marguerite, terre Graham, (Lat. 67°53'20”S ; Long. 67°25'30”O), à 65 km à l'est (Carte 1).
6 (v) Zones spéciales à l'intérieur de la zone
Aucune.
7. Critères de délivrance des permis
7 (i) Conditions générales pour l'obtention d'un permis
L'accès à la zone est interdit sauf avec un permis délivré par une autorité nationale compétente. Les critères de délivrance d'un permis pour entrer dans la zone sont les suivants :
- un permis est délivré pour des raisons scientifiques indispensables, qu'il est impossible de satisfaire ailleurs ou pour des raisons de gestion essentielles à la zone ;
- les activités autorisées sont conformes au présent plan de gestion ;
- toutes les activités de gestion soutiennent la réalisation des buts et objectifs du présent plan de gestion ;
- les activités autorisées ne mettront pas en péril l'écosystème naturel de la zone ;
- les activités autorisées veilleront, au moyen d'un processus d'évaluation d'impact sur l'environnement, à la protection permanente des valeurs environnementales ou scientifiques de la zone ;
- le permis est délivré pour une durée déterminée ; et
- la détention du permis ou d'une copie certifiée conforme est impérative dans la zone.
7 (ii) Accès à la zone et déplacements à l'intérieur de celle-ci
L'utilisation de véhicules terrestres (motoneiges, quads, etc.) dans la zone est interdite.
Tout déplacement sur la terre ferme dans la zone doit être effectué à pied. Les déplacements à pied doivent être limités au minimum requis pour effectuer les activités autorisées et tous les efforts raisonnables doivent être consentis pour réduire les effets du piétinement.
Les piétons respecteront les itinéraires tracés afin de réduire au maximum les perturbations des oiseaux en phase de reproduction et emprunteront au besoin un itinéraire plus long pour parvenir au lieu de destination.
Les itinéraires de marche ont été conçus pour éviter les sites de reproduction les plus sensibles et ils doivent être utilisés lorsqu'il est indispensable de traverser l'île (Carte 2). Les visiteurs ne doivent jamais perdre de vue que les sites de nidification spécifiques peuvent varier d'année en année et que, par conséquent, le tracé de l'itinéraire recommandé peut changer. Les itinéraires sont proposés à titre indicatif et les visiteurs sont appelés à faire preuve de discernement pour limiter les effets de leur présence. Dans certaines parties de l'île, lorsque cela est possible et sûr, il est souvent préférable d'opter pour un itinéraire qui suit le littoral de la zone. Trois itinéraires sont désignés (Carte 2) : l'itinéraire n° 1 traverse la partie centrale de l'île et relie les refuges chilien et argentin. L'itinéraire n° 2 facilite l'accès aux phares sur le sud de l'île et s'étend du centre de la côte est jusqu'aux versants orientaux de la colline. Cependant, lors d'une visite de gestion en 2011, cet itinéraire s'est révélé être colonisé par des oiseaux. C'est pourquoi l'itinéraire n° 3 a également été conçu. Il fait route vers l'est à partir du refuge argentin jusqu'à un passage étroit sur le côté ouest de l'île, et continue au sud-ouest jusqu'à un ravin/versant, vers une surface plane au-dessus des colonies abandonnées (depuis janvier 2011) de cormorans impériaux. A partir de là, l'itinéraire emprunte une trajectoire vers l'est en direction des phares. Des précautions doivent être prises pour éviter de piétiner les bancs de mousse qui se trouvent à proximité d'un bassin d'eau de fonte, à environ 70 mètres au nord des phares.
L'accès aux endroits privilégiés par les pétrels géants pour la nidification (Carte 3) sera uniquement autorisé dans le cadre des objectifs spécifiés dans le permis. Lorsqu'il est indispensable d'accéder au phare (pour procéder à son entretien par exemple), les visiteurs devront suivre l'itinéraire prévu le plus rigoureusement possible en veillant à éviter les oiseaux nicheurs. La majeure partie de la zone menant au phare et entourant ce dernier est occupée par des pétrels en phase de reproduction. Une grande prudence est donc de rigueur.
Les mouvements doivent être lents, silencieux et toujours à bonne distance des oiseaux nicheurs.
Les visiteurs seront particulièrement attentifs aux signes d'agitation et feront même demi-tour en cas de forte perturbation.
Les opérations de survol de la zone doivent être réalisées conformément aux Directives pour l'exploitation d'aéronefs à proximité de concentrations d'oiseaux dans l'Antarctique, inscrites dans la résolution 2 (2004).
Le survol de colonies d'oiseaux dans la zone par des systèmes d'aéronefs pilotés à distance (RPAS) n'est pas autorisé, sauf à des fins scientifiques ou opérationnelles, et en vertu d'un permis émis par une autorité nationale compétente.
7 (iii) Activités pouvant être conduites à l'intérieur de la zone
Les activités pouvant être menées sont :
- les activités de gestion essentielles, y compris de suivi ;
- des travaux de recherche scientifique indispensables qui ne peuvent être entrepris ailleurs et ne risquent pas de mettre en péril l'écosystème de la zone ; et
- l'échantillonnage, qui doit être réduit au minimum pour répondre aux programmes de recherches approuvés.
Les restrictions concernant les périodes autorisées pour mener des activités s'appliquent à la zone et sont spécifiées dans les sections pertinentes du présent plan de gestion.
7 (iv) Ins tallation, modification ou enlèvement de structures
Toute nouvelle structure ou installation permanente est interdite dans la zone.
Les structures existantes qui sont désaffectées ou saccagées doivent être enlevées ou rénovées.
Toute activité liée à l'installation, à la modification, à l'entretien ou à l'enlèvement de structures doit être réalisée de manière à minimiser les perturbations des oiseaux en phase de reproduction. Elle devra avoir lieu entre le 1er février et le 30 septembre inclus pour éviter la grande saison de reproduction.
Aucune structure ne peut être érigée dans la zone et aucun matériel scientifique ne peut y être installé, sauf pour des raisons scientifiques ou de gestion indispensables et pour une période préétablie définies dans un permis.
Toutes les bornes, les structures et tout l'équipement scientifique installés dans la zone doivent clairement identifier le pays, le nom du principal chercheur ou de la principale agence, l'année d'installation et la date d'enlèvement prévue.
Tous ces objets doivent être exempts de tout organisme, propagule (semence, œufs, spores) ou terre non stérile (voir section 7 (vi)), et doivent être composés de matériaux résistants aux conditions environnementales et présenter un risque de contamination minime pour la zone.
L'enlèvement d'équipements spécifiques pour lesquels le permis a expiré est du ressort de l'autorité qui a délivré le permis d'origine et il sera l'un des critères régissant la délivrance du permis.
7 (v) Emplacement des camps
Tout campement doit être évité dans la zone. Cependant, lorsque certaines opérations autorisées par le permis l'exigent, des camps temporaires peuvent être installés à deux endroits désignés : sur la côte est centrale de l'île (Lat. 67°46'25.8”S ; Long. 68°53'00.8”O) et sur la côte nord-ouest centrale de la zone (Lat. 67°46'08.2”S ; Long. 68°53'29.5”O) (Carte 2).
7 (vi) Restrictions relatives aux matériaux et organismes pouvant être introduits dans la zone
L'introduction délibérée dans la zone d'animaux, de végétaux ou de micro-organismes vivants est interdite. Pour garantir le maintien des valeurs floristiques et écologiques de la zone, des précautions spéciales devront être prises pour prévenir toute introduction accidentelle de microbes, d'invertébrés ou de plantes provenant d'autres sites antarctiques, y compris des stations, ou d'autres régions que l'Antarctique. Tout le matériel d'échantillonnage et les balises introduits dans la zone doivent être nettoyés et stérilisés. Dans la mesure du possible, les chaussures et autres équipements utilisés ou introduits dans la zone (y compris les sacs et les sacs à dos) doivent être minutieusement nettoyés avant d'entrer dans la zone. Le Manuel sur les espèces non indigènes du CPE (CPE, 2017) et le « Environmental code of conduct for terrestrial scientific field research in Antarctica » [Code de conduite environnemental pour la recherche scientifique sur le terrain en Antarctique] (SCAR, 2009) offrent des orientations supplémentaires en la matière. Compte tenu de la présence de colonies d'oiseaux nicheurs dans la zone, aucun produit provenant ou dérivé d'espèces avicoles - notamment les déchets et les produits contenant des œufs en poudre crus - ne doit être introduit dans la zone ou dans sa composante marine.
Aucun herbicide ni pesticide ne doit être introduit dans la zone. Tous autres produits chimiques, y compris les radionucléides ou les isotopes stables, qui peuvent être introduits pour des raisons scientifiques ou des raisons de gestion visées dans le permis, seront enlevés de la zone au plus tard à la fin de l'activité pour laquelle le permis a été délivré. L'émission directe de radionucléides ou d'isotopes stables dans l'environnement par une méthode qui les rendrait irrécupérables est à éviter. Les carburants et autres produits chimiques ne doivent pas être entreposés dans la zone, hormis sur délivrance d'un permis. Auquel cas, ces matériaux doivent être entreposés et manipulés de façon à limiter les risques d'introduction accidentelle dans l'environnement. Tous les matériaux sont introduits dans la zone pour une période déterminée uniquement ; ils doivent être enlevés lorsque cette période est échue. En cas d'introduction susceptible de porter préjudice aux valeurs de la zone, les matériaux seront retirés dans la mesure où ce retrait n'entraine pas de conséquences plus graves que de les laisser in situ. L'autorité compétente doit être notifiée de tout élément libéré ou émis dans la zone et qui n'en a pas enlevé, à moins que cela soit autorisé par le permis.
7 (vii) Prélèvements de végétaux, capture d'animaux ou perturbations nuisibles de la faune et la flore
Toute capture d'animaux ou toute perturbation nuisible à la faune et la flore indigène est interdite sauf avec un permis distinct délivré spécifiquement à cette fin en vertu de l'annexe II du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement. Dans le cas de prélèvements ou de perturbations nuisibles d'animaux, il convient d'appliquer comme norme minimale le « SCAR code of conduct for the use of animals for scientific purposes in Antarctica » [Code de conduite du SCAR pour l'utilisation d'animaux à des fins scientifiques dans l'Antarctique] (2011). Les activités d'échantillonnage des sols et de la végétation doivent être limitées au minimum absolu requis à des fins scientifiques ou à des fins de gestion et exécutées avec des techniques qui réduisent au maximum les perturbations susceptibles d'être causées au sol et au biote environnants.
7 (viii) Collecte ou retrait de matériaux non introduits dans la zone par le titulaire du permis
Les matériaux ne peuvent être ramassés ou enlevés de la zone qu'en conformité avec un permis et doivent être limités au minimum requis pour répondre aux besoins scientifiques ou de gestion. Les matériaux d'origine humaine susceptibles de mettre en péril les valeurs de la zone, qui n'ont pas été introduits dans celle-ci par le détenteur du permis ou qui n'ont pas été autrement autorisés, peuvent être enlevés de la zone à moins que l'impact environnemental de l'enlèvement ne soit plus grand que si les matériaux sont laissés in situ. Si tel est le cas, l'autorité compétente doit en être informée et son autorisation obtenue. Un permis ne sera pas délivré s'il s'avère à juste titre que l'échantillonnage proposé prélèverait, enlèverait ou endommagerait de telles quantités de substrat et de faune et de flore sauvages que leur répartition ou leur abondance sur l'île serait sérieusement affectée. Les échantillons de faune et de flore retrouvés sans vie dans la zone peuvent être enlevés à des fins d'analyse et de vérification sans autorisation préalable.
7 (ix) Elimination des déchets
Tous les déchets, à l'exception des déchets humains, seront retirés de la zone. Dans l'idéal, tous les déchets humains devraient être ôtés de la zone. Si cela n'est pas possible, ils peuvent être jetés à la mer.
7 (x) Mesures qui peuvent être nécessaires pour continuer de répondre aux objectifs du plan de gestion
1. Des permis peuvent être délivrés pour entrer dans la zone afin d'y faire des travaux de recherche scientifique, de surveillance et d'inspection de site, qui peuvent impliquer le prélèvement d'un petit nombre d'échantillons à des fins d'analyse, ou pour appliquer des mesures de protection.
2. Tous les sites de suivi de longue durée doivent être signalés par des bornes ou des panneaux dûment entretenus.
3. Les activités de nature scientifique seront menées conformément au « Environmental code of conduct for terrestrial scientific field research in Antarctica » [Code de conduite environnemental pour la recherche scientifique sur le terrain en Antarctique] (SCAR, 2009).
7 (xi) Rapports de visite
Le principal détenteur du permis soumettra, pour chaque visite dans la zone, un rapport à l'autorité nationale compétente, dès que possible, et au plus tard six mois après la fin de ladite visite. Ces rapports doivent inclure, s'il y a lieu, les renseignements identifiés dans le formulaire de rapport de visite des zones spécialement protégées de l'Antarctique qui figure dans le Guide pour la préparation des plans de gestion des zones spécialement protégées de l'Antarctique (annexe 2). Les autorités compétentes doivent être informées de toute activité ou mesure qui ne serait pas autorisée par le permis. Si cela est possible, l'autorité nationale doit également transmettre une copie du rapport de visite à la Partie qui a proposé le plan de gestion et ce, afin d'aider à gérer la zone et à revoir le plan de gestion. Les Parties doivent, dans la mesure du possible, déposer les originaux ou les copies de ces rapports dans une archive à laquelle le public pourra avoir accès afin de conserver une archive d'usage qui sera utilisée pour toute révision du plan de gestion et pour l'organisation de l'utilisation scientifique de la zone.
8. Bibliographie
Barlow, J. (1968). Biological report. Adelaide Island. 1967/68. Rapport non publié de la British Antarctic Survey, Archives de la BAS Réf. AD6/2T/1967/N.
Bramwell, M. J. (1969). Report on elephant seal pupping on Avian Island. Rapport non publié de la British Antarctic Survey, Archives de la BAS Réf. AD6/2T/1969/N.
Bramwell, M. J. (1970). Journey report : Avian Island 7 Oct - 4 Nov 1969. Rapport non publié de la British Antarctic Survey, Archives de la BAS Réf. AD6/2T/1969/K3.
Casanovas, P., Naveen, R., Forrest, S., Poncet, J. & Lynch, H.J. (2015). A comprehensive coastal seabird survey maps out the front lines of ecological change on the western Antarctic Peninsula. Polar Biology 38 : 927-940.
Comité pour la protection de l'environnement (CPE). (2017). Manuel sur les espèces non indigènes - 2e édition. Manuel préparé par le Groupe de contact intersessions du CPE et adopté par la Réunion Consultative du Traité sur l'Antarctique dans la résolution 4 (2016). Buenos Aires, Secrétariat du Traité sur l'Antarctique.
Elliott, M. H. (1969). Summer geological camp on Avian Island 26 Nov - 4 Dec 1968. Rapport non publié de la British Antarctic Survey, Archives de la BAS Réf. AD6/2T/1968/K3.
Fox, A., & Gray, M. (1997). Aerial photography field report 1996-97 Antarctic field season. Rapport non publié de la British Antarctic Survey, Archives de la BAS Réf. AD6/2R/1996/L2.
Gray, M. , & Fox, A. (1997). GPS Survey field report 1996-97 Antarctic field season. Rapport non publié de la British Antarctic Survey, Archives de la BAS Réf. AD6/2R/1996/L1.
Griffiths, C. (1992). Geological fieldwork on Adelaide Island 1991-92. Rapport non publié de la British Antarctic Survey, Archives de la BAS Réf. AD6/2R/1991/GL1.
Harris, C.M. (2001). Revision of management plans for Antarctic protected areas originally proposed by the United States of America and the United Kingdom : Field visit report. Rapport interne pour la Fondation nationale pour la science des États-Unis et le Bureau des Affaires étrangères et du Commonwealth du Royaume-Uni. Environmental Research & Assessment, Cambridge.
Harris, C.M., Lorenz, K., Fishpool, L.D.C., Lascelles, B., Cooper, J., Coria, N.R., Croxall, J.P., Emmerson, L.M., Fijn, R.C., Fraser, W.L., Jouventin, P., LaRue, M. A., Le Maho, Y., Lynch, H.J., Naveen, R., Patterson- Fraser, D.L., Peter, H.-U., Poncet, S., Phillips, R.A., Southwell, C.J., van Franeker, J.A., Weimerskirch, H., Wienecke, B., & Woehler, E.J. (2015). zones importantes pour la conservation des oiseaux de l'Antarctique 2015. BirdLife International et Environmental Research & Assessment Ltd., Cambridge.
Lynch, H. J., Naveen, R., Trathan, P. N., & Fagan, W. F. (2012). Spatially integrated assessment reveals widespread changes in penguin populations on the Antarctic Peninsula. Ecology 93 : 1367-1377.
Moyes, A. B., Willan, C. F. H., Thomson, J. W., et al. (1994). Geological map of Adelaide Island to Foyn Coast, BAS GEOMAP Series, Sheet 3, Scale 1 : 250,000, with supplementary text. British Antarctic Survey, Cambridge.
Patterson, D. L., Woehler, E. J., Croxall, J. P., Cooper, J., Poncet, S., Peter, H.-U., Hunter, S., and Fraser, W.
R. (2008). Breeding distribution and population status of the northern giant petrel Macronectes halli and the southern giant petrel M. giganteus. Marine Ornithology 36 : 115-124.
Poncet, S., & Poncet, J. (1979). Ornithological report, Avian Island, 1978-79. Rapport non publié de la British Antarctic Survey, Archives de la BAS Réf. AD6/2R/1978/Q.
Poncet, S. (1982). Le grand hiver : Damien II base Antarctique. Les Éditions Arthaud, Paris.
Poncet, S., & Poncet, J. (1987). Censuses of penguin populations of the Antarctic Peninsula, 1983-87. Bulletin n° 77 de la British Antarctic Survey : 109-129.
Poncet, S. (1990). Avian Island, Marguerite Bay, Antarctic Peninsula, SPA proposal. Rapport non publié du Groupe d'experts du SCAR sur l'environnement et la conservation, 1990.
Riley, T. R., Flowerdew, M. J. & Whitehouse, M. J. (2012). Litho- and chronostratigraphy of a fore- to intra- arc basin : Adelaide Island, Antarctic Peninsula. Geological Magazine 149 : 768-782.
Ritz, M. S., Hahn, S., Janicke, T., & Peter, H.-U. (2006). Hybridisation between South Polar Skua (Catharacta maccormicki) and Brown Skua (C. antarctica lonnbergi) in the Antarctic Peninsula region. Polar Biology 29 : 153-159.
Sailley, S. F., Ducklow, H. W., Moeller, H. V., Fraser, W. R., Schofield, O. M. , Steinberg, D. K., Price, L. M. , & Doney, S. C. (2013). Carbon fluxes and pelagic ecosystem dynamics near two western Antarctic Peninsula Adélie penguin colonies : an inverse model approach. Marine Ecology Progress Series 492 : 253- 272.
SCAR (Comité scientifique pour la recherche antarctique). (2009). Environmental code of conduct for terrestrial scientific field research in Antarctica [Code de conduite environnemental pour la recherche scientifique sur le terrain en Antarctique]. XXXIIe RCTA IP4.
SCAR (Comité scientifique pour la recherche antarctique). (2011). SCAR code of conduct for the use of animals for scientific purposes in Antarctica [Code de conduite du SCAR pour l'utilisation d'animaux à des fins scientifiques]. XXXIVe RCTA IP53.
Smith, H. G. 1978. The distribution and ecology of terrestrial protozoa of sub-Antarctic and maritime Antarctic islands. Rapport scientifique n° 95 du BAS, British Antarctic Survey, Cambridge.
Smith, R. I. L. (1996). Terrestrial and freshwater biotic components of the western Antarctic Peninsula, dans Ross, R.M., Hofmann, E.E. et Quetin, L.B. Foundations for ecological research west of theAntarctic Peninsula. Antarctic Research Series 70 : American Geophysical Union, Washington D.C. : 15-59.
Stonehouse, B. (1949). Report on biological activities at Base E 1948-49. Rapport non publié de la British Antarctic Survey, Archives de la BAS Réf. AD6/2E/1948/N1.
Stonehouse, B. (1950). Preliminary report on biological work Base E 1949-50. Rapport non publié de la British Antarctic Survey, Archives de la BAS Réf. AD6/2E/1949/N.
Terauds, A., & Lee, J. R. (2016). Antarctic biogeography revisited : updating the Antarctic Conservation Biogeographic Regions. Diversity and Distribution 22 : 836-840.
Terauds, A., Chown, S. L., Morgan, F., Peat, H. J., Watt, D., Keys, H., Convey, P., & Bergstrom, D. M. (2012). Conservation biogeography of the Antarctic. Diversity and Distributions 18 : 726-41.
Willey, I. M. (1969). Adelaide Island bird report 1968. Rapport non publié de la British Antarctic Survey, Archives de la BAS Réf. AD6/2T/1968/Q.
Woehler, E. J. (ed). (1993). The distribution and abundance of Antarctic and sub-Antarctic penguins. SCAR, Cambridge.
Carte 1. - Ile Avian, ZSPA N° 117, par rapport à la baie Marguerite, indiquant l'emplacement des stations Teniente Luis Carvajal (Chili), Rothera (RU) et General San Martín (Argentine). Emplacements d'autres zones protégées au sein de la baie Marguerite (ZSPA n° 107 sur l'île Emperor (îles Dion), ZSPA n° 115 sur l'île Lagotellerie et ZSPA n° 129 sur la pointe Rothera) sont également indiqués. Encart : emplacement de l'île Avian sur la péninsule antarctique.
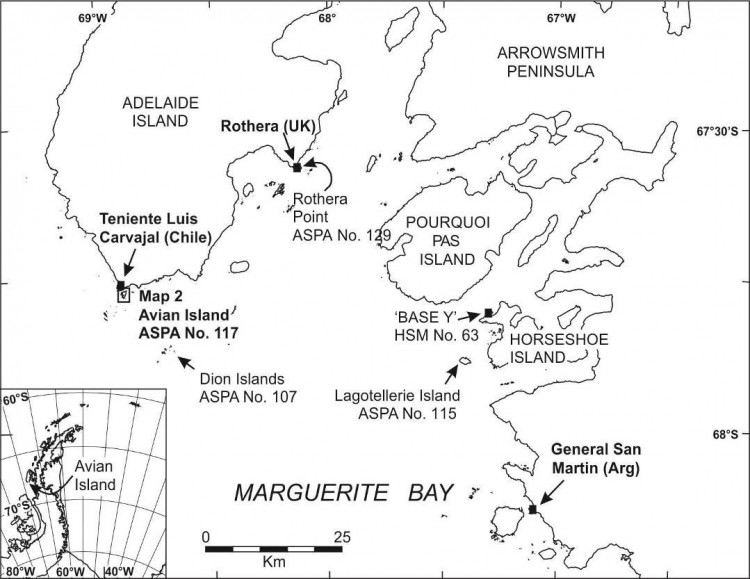
Carte 2. - Ile Avian (ZSPA n° 117), carte topographique. Spécifications de la carte - projection : conique conforme de Lambert ; parallèles types : 1er 67°30'00"S ; 2e 68°00'00”S ; méridien central : 68°55'00”O ; latitude d'origine : 68°00'00”S ; sphéroïde : WGS84 ; datum : niveau moyen de la mer ; équidistance des courbes de niveau verticales : 5 mètres ; précision horizontale : ± 5 mètres ; précision verticale ±1,5 mètres.
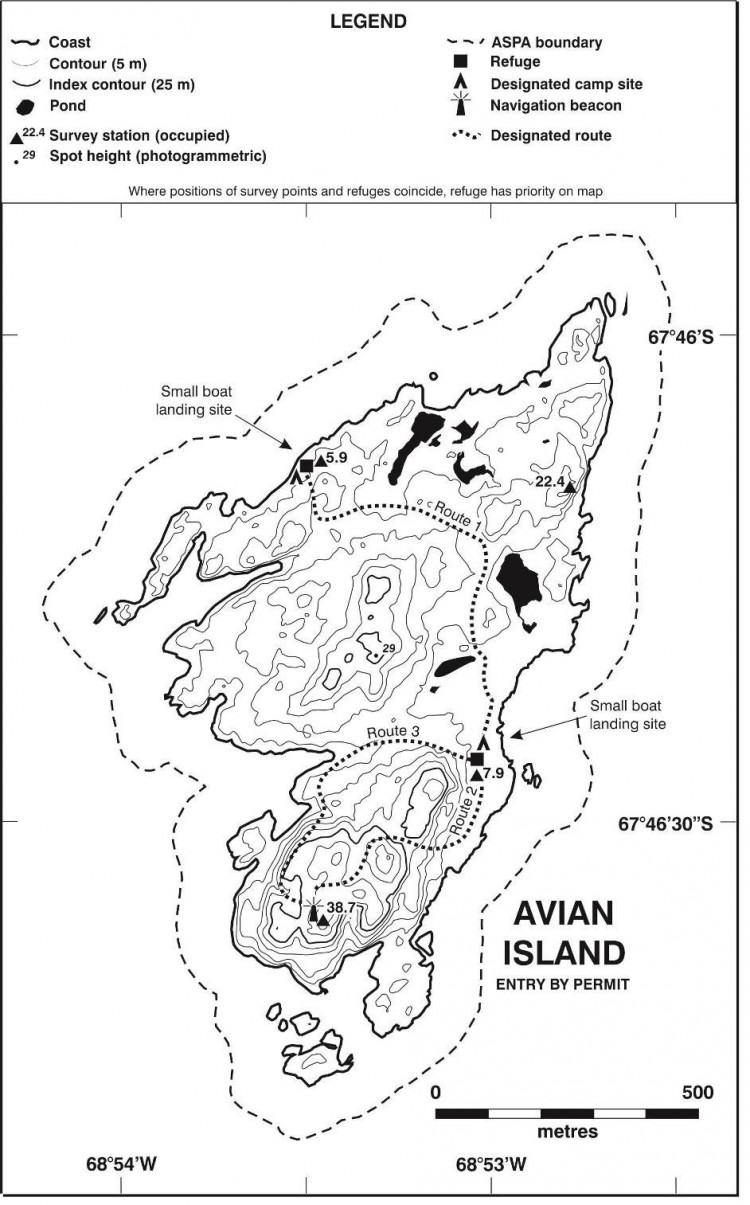
Carte 3. - Ile Avian (ZSPA n° 117), croquis cartographique de la faune en phase de reproduction. La position des nids et des colonies sont d'une précision de ± 25 m. Ces informations proviennent de Poncet (1982). Spécifications de la carte - projection : conique conforme de Lambert ; parallèles types : 1er 67°30'00”S ; 2e 68°00'00”S ; méridien central : 68°55'00”O ; latitude d'origine : 68°00'00”S ; sphéroïde : WGS84 ; datum : niveau moyen de la mer ; équidistance des courbes de niveau verticales : 5 mètres ; précision horizontale : ± 5 mètres ; précision verticale ± 1,5 mètres.
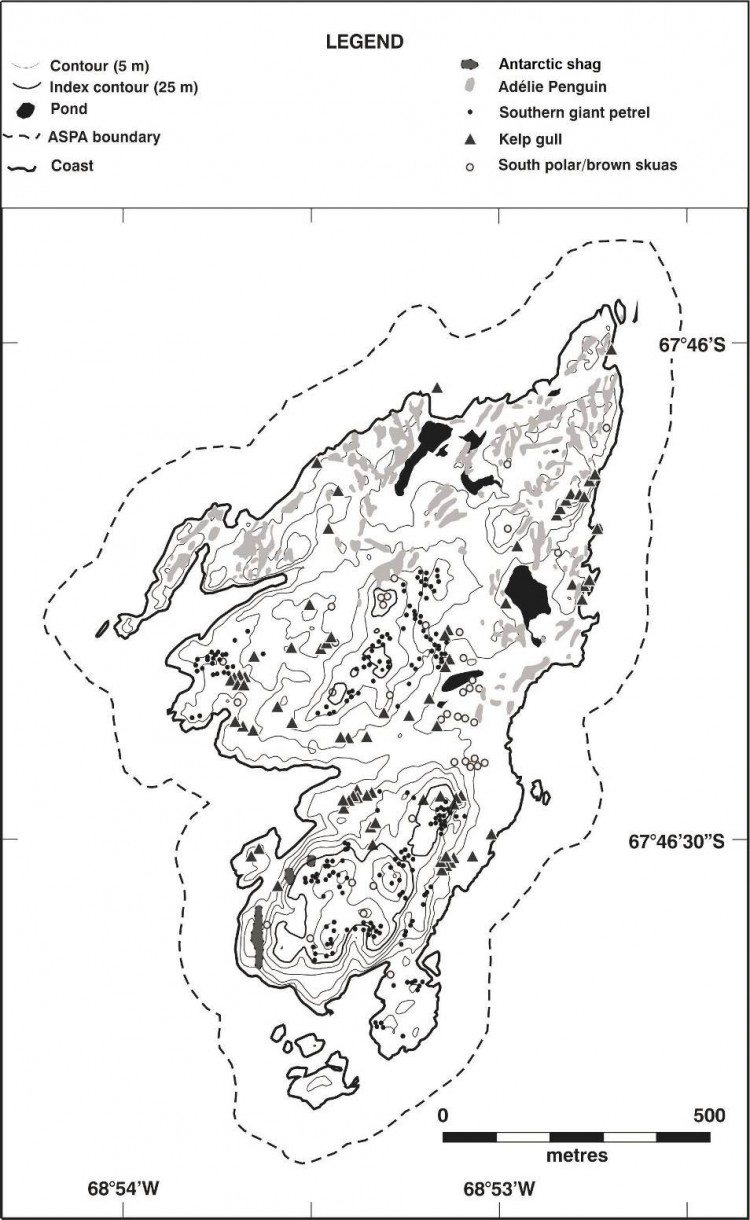
Annexe
MESURE 3 (2018)
ZONE SPÉCIALEMENT PROTÉGÉE DE L'ANTARCTIQUE N° 132 (PÉNINSULE POTTER, ÎLE DU ROI-GEORGE [ISLA 25 DE MAYO], ÎLES SHETLAND DU SUD) : PLAN DE GESTION RÉVISÉ (ENSEMBLE UNE ANNEXE)
Les Représentants,
Rappelant les articles 3, 5 et 6 de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement, qui disposent de la désignation des zones spécialement protégées de l'Antarctique (« ZSPA ») et de l'adoption des plans de gestion pour ces zones ;
Rappelant :
- la recommandation XIII-8 (1985), qui désignait la péninsule Potter, île du Roi-George (Isla de 25 Mayo), îles Shetland du Sud comme site présentant un intérêt scientifique particulier (« SISP ») n° 13 et annexait un plan de gestion pour le site ;
- la mesure 3 (1997), qui a mis en annexe un plan de gestion révisé pour le SISP n° 13 ;
- la décision 1 (2002), qui a renommé et renuméroté le SISP n° 13 en ZSPA n° 132 ;
- la mesure 2 (2005) et la mesure 4 (2013), qui ont adopté des plans de gestion révisés pour la ZSPA n° 132 ;
Rappelant que la mesure 3 (1997) n'est pas encore entrée en vigueur ;
Notant que le Comité pour la protection de l'environnement a approuvé un plan de gestion révisé pour la ZSPA n° 132 ;
Souhaitant remplacer le plan de gestion actuel pour la ZSPA n° 132 avec le plan de gestion révisé ;
Recommandent à leurs gouvernements d'approuver la mesure suivante conformément au paragraphe 1 de l'article 6 de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement :
Que :
1. Le plan de gestion révisé pour la zone spécialement protégée de l'Antarctique n° 132 (péninsule Potter, île du Roi-George [Isla 25 de Mayo], îles Shetland du Sud), qui figure en annexe de cette mesure, soit approuvé ; et que
2. Le plan de gestion pour la zone spécialement protégée de l'Antarctique n° 132 annexé à la mesure 4 (2013) soit abrogé.
Annexe
PLAN DE GESTION POUR LA ZONE SPÉCIALEMENT PROTÉGÉE DE L'ANTARCTIQUE (ZSPA) Nº 132
PÉNINSULE POTTER
Introduction
A l'origine, cette zone a été désignée comme Site d'intérêt scientifique particulier n° 13 (recommandation XIII-8, XIIIe RCTA, Bruxelles, 1985) sur proposition de l'Argentine, compte tenu de la diversité et de l'étendue de sa végétation et de sa faune qui constituent un échantillon représentatif de l'écosystème antarctique.
En 1997, le plan de gestion a été adapté aux critères de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement et approuvé par le biais de la Mesure 3 (1997). Cette version correspond à la révision du plan de gestion approuvé conformément à la Mesure 2 (2005) et il s'agit de la seconde révision depuis l'entrée en vigueur de l'annexe V.
Les objectifs initiaux de désignation de cette zone demeurent valides. La péninsule Potter a été désignée comme zone spécialement protégée de l'Antarctique pour protéger ses valeurs environnementales exceptionnelles et pour faciliter les travaux de recherche scientifique en cours et à venir. Les perturbations anthropiques pourraient porter atteinte aux études de longue durée qui y sont menées, en particulier pendant les périodes de reproduction, ou affecter les niveaux de référence des polluants chimiques dans les matrices biotiques et/ou abiotiques (par exemple éléments trace et composés organiques persistants).
La raison principale de la désignation de la ZSPA est que la péninsule Potter constitue un échantillon représentatif de groupements d'espèces de l'écosystème antarctique. Ses régions côtières abritent d'importantes colonies d'oiseaux, des sites de reproduction de mammifères marins et diverses espèces végétales. Ces côtes sont actuellement les plus sensibles aux changements climatiques et à ses effets indirects, comme la fonte des glaces (Hernando et al. (2015) qui a des effets prouvés sur la biodiversité (Sahade et al. 2015). Elle présente donc une valeur scientifique considérable puisque de nombreuses études peuvent y être menées sur les impacts du changement climatique dans les facteurs biotiques et abiotiques ainsi que ses conséquences sur la chaîne alimentaire (p. ex. Carlini et al. 2009, Carlini et al. 2010, Casaux et al. 2006, Daneri et Carlini 1999, Rombolá et al. 2010, Torres et al. 2012, Quillfeldt et al. 2017, Juáres et al., 2018). Il est capital de maintenir ces activités scientifiques, telles que le programme de surveillance qui y est mené depuis 1982, comprenant le Programme de contrôle de l'écosystème de la CCAMLR (CEMP), initié en 1995, pour les données scientifiques inestimables que celui-ci peut apporter. De plus, les connaissances sur la dynamique des planctons (Bers et al., Schloss et al. 2014) et du krill (Di Fonzo et al. 2014, 2017a, 2017b, Fuentes et al. 2016), qui constituent la base alimentaire des organismes plus imposants dans le réseau trophique, sont d'une importance cruciale.
Plusieurs caractéristiques rendent cette zone particulièrement sensible aux perturbations humaines, comme sa configuration, à savoir une zone littorale relativement étroite, comprise entre la mer et une falaise, où il existe très peu d'espace d'opération sans interférence avec les colonies en phase de reproduction. La forte concentration d'activités, de stations scientifiques, et l'accessibilité facile de la zone par voie maritime ou terrestre, même si elle implique de petites embarcations, constituent une menace potentielle pour les valeurs biologiques et les activités de recherche.
D'après de récentes études, la situation environnementale dans les îles Shetland du Sud montre que le détroit de Bransfield, dans la partie de l'océan Austral proche de la péninsule Potter, a été sérieusement transformé, tout d'abord par la disparition presque totale de la population autrefois incroyablement abondante d'otaries à fourrure (Arctocephalus spp.) se nourrissant de poissons et de krill, suivie de celle des mysticètes. Plus récemment, les otaries à fourrure se sont largement rétablies et les mysticètes commencent à prendre cette voie (Ainley et al., 2010). Toutefois les changements climatiques modifient de plus en plus les processus écologiques, ce qui se traduit par des changements physiques dans les températures, la circulation d'eau et l'expansion de la glace marine, entre autres. Suite à la réduction du nombre de proies, non seulement du fait des changements climatiques et du rétablissement des espèces concurrentes, mais également en raison d'autres facteurs actuellement inconnus, les populations de manchots diminuent (Ducklow et al. 2007, Ainley et Blight 2009, Ainley et al. 2010, Trivelpiece et al. 2011, Juáres et al. 2015). En ce sens, actuellement, la ZSPA n° 132 présente aujourd'hui une pertinence particulière, étant donné que l'étude des colonies de manchots Adélie qui se trouvent dans la zone apporte des réponses quant aux changements environnementaux observés dans la péninsule antarctique, notamment la fréquence moins élevée des années froides associée à la réduction des étendues de glace marine, et ses impacts sur l'abondance du krill. (Garcia et al. 2015). Elle permet aussi de détecter et d'enregistrer des changements importants dans l'écosystème marin et vise à faire la différence entre les changements causés par le prélèvement commercial d'espèces et ceux causés par des variables environnementales, qu'elles soient physiques ou biologiques.
La péninsule Potter offre également des opportunités exceptionnelles pour la réalisation d'autres études scientifiques portant sur les communautés biologiques terrestres et marines.
Les recherches et les programmes de surveillance actuellement menés dans la ZSPA n° 132 incluent notamment :
- Biosurveillance côtière : impact des changements climatiques mondiaux et des xénobiotiques sur les espèces clés des réseaux trophiques antarctiques ;
- Polluants organiques persistants et éléments traces dans les matrices biotiques et abiotiques de l'environnement antarctique ;
- Création d'énergie, types de proies et réponses éventuelles des pinnipèdes aux anomalies climatiques et à l'extension de glace marine dans la région de la péninsule antarctique et de l'arc de Scotia ;
- Réponse des populations d'oiseaux de l'Antarctique à la variabilité interannuelle de leurs proies dans des zones où les impacts du changement climatique sont évidents ;
- Phylogéographie de la Deschampsia Antarctica basée sur des études moléculaires, morphologiques et caryologiques ;
- Répartition et état nutritionnel des labbes antarctiques et des labbes de McCormick ;
- Programme de contrôle de l'écosystème de la CCAMLR - Site du CEMP depuis 1995.
1. Description des valeurs à protéger
Les régions littorales abritent d'importantes colonies d'oiseaux, des colonies de mammifères marins en phase de reproduction et une végétation abondante (vastes tapis de mousses et de lichens, touffes de canche antarctique et de tillandsia [Deschampsia antarctica and Colobanthus quitensis] dans les zones côtières). Des programmes de recherche scientifique sur l'écologie de la reproduction d'espèces d'oiseaux et de mammifères marins (comme les éléphants de mer [Mirounga leonina], des manchots d'Adélie [Pygoscelis adeliae] et des manchots papous [Pygoscelis papua] ) comprenant entre autres le Programme de contrôle de l'écosystème de la CCAMLR, sont mis en place dans la zone depuis 1982. Les colonies en phase de reproduction se trouvent dans un lieu spécifique sur la côte. La zone comprend principalement des plages surélevées, en grande partie recouvertes de pierres de taille moyenne, de structures basaltiques et de moraines latérales et terminales. Le littoral est très irrégulier et se compose d'une série de petites baies formées parmi des promontoires rocheux, où l'on trouve fréquemment plusieurs espèces de pinnipèdes de l'Antarctique. Ils viennent généralement dans cette zone pour se reproduire ou pour muer. Les raisons susmentionnées confèrent à la zone des valeurs scientifiques et esthétiques exceptionnelles.
Bien que l'Antarctique soit considéré comme l'une des rares régions non polluées de la planète, du fait de son relatif isolement par rapport aux centres industriels et urbains, certaines études révèlent des aires polluées près des stations scientifiques. La station Carlini avoisinante est notamment concernée (Curtosi et al. 2010, Vodopivez et al. 2015). Il apparaît donc nécessaire de redoubler de vigilance au sein de la ZSPA n° 132.
D'après Morgan et al. (2007), la ZSPA n° 132 représente le domaine environnemental " Îles au large des côtes de la péninsule antarctique ". En outre, d'après Terauds et al. (2012), la péninsule Potter représente la région " Nord-Est de la péninsule antarctique "selon la classification des Régions de conservation biogéographiques de l'Antarctique et la zone 047 des " zones importantes pour la conservation des oiseaux 2015 "(Harris et al., 2015).
Pour des caractéristiques plus détaillées, il convient de se reporter à la section 6.
2. Buts et objectifs
Préserver l'écosystème naturel et éviter les perturbations humaines inutiles.
Permettre la réalisation de travaux de recherche scientifique sous réserve qu'ils ne portent pas atteinte aux valeurs de la zone.
Eviter tout changement important dans la structure et la composition des communautés faunistiques et floristiques.
Protéger la flore de la zone des impacts anthropiques, pour qu'elle serve de référence.
Eviter ou réduire au maximum l'introduction de plantes, d'animaux et de microbes non indigènes dans la zone.
Limiter le risque d'introduction d'agents pathogènes qui pourraient provoquer des maladies au sein des populations fauniques de la zone.
Eviter l'introduction, la production ou la diffusion de polluants chimiques pouvant affecter la zone.
3. Activités de gestion
Le personnel affecté à la base Carlini (anciennement base Jubany, base argentine adjacente à la ZSPA), en particulier le personnel autorisé à entrer dans la ZSPA, recevra une formation particulière relative aux conditions qui régissent le plan de gestion.
Des copies de ce plan de gestion doivent être disponibles à la base Carlini.
Des distances d'approche maximales de la faune doivent être respectées, sauf si les projets scientifiques ont d'autres exigences, et sous réserve que les permis adéquats aient été délivrés.
Le prélèvement d'échantillons est limité au strict nécessaire à la mise en œuvre de programmes de recherche scientifique autorisés.
Tous les repères et toutes les structures érigés dans la ZSPA à des fins scientifiques ou de gestion doivent être solidement fixés et soigneusement entretenus.
Conformément aux dispositions de l'annexe III du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement, les équipements ou matériels abandonnés seront retirés dans toute la mesure du possible, à condition que leur retrait n'ait pas d'impact préjudiciable sur l'environnement et les valeurs de la zone.
Le plan de gestion sera réexaminé au moins tous les 5 ans et mis à jour en conséquence.
Tous les pilotes qui opèrent dans la région seront tenus informés de l'emplacement de la zone, des limites de celle-ci et des restrictions qui s'appliquent à l'entrée et au survol dans la zone.
4. Durée de désignation
La zone est désignée pour une période indéterminée.
5. Cartes et photographies
La Carte 1, comprise à la fin du présent plan de gestion, montre l'emplacement de la ZSPA n° 132 (lignes diagonales) par rapport à la péninsule Potter, île du Roi-George (Isla 25 de Mayo).
6. Description de la zone
6 (i) Coordonnées géographiques, bornage et caractéristiques du milieu naturel
Coordonnées géographiques et limites
La zone est située sur la côte de la baie Maxwell, au sud-est de l'île du Roi-George (Isla 25 de Mayo), entre l'extrême sud de la pointe Mirounga (au nord-est de la péninsule Potter) et l'affleurement rocheux connu sous le nom de " Saillie 7 ", sur la limite nord-est de la pointe Stranger. La zone s'étend le long de la bande littorale, à marée basse, jusqu'au bord de la falaise, atteignant des hauteurs de 15 - 50 mètres. La façade du bord de la falaise est incluse dans la ZSPA. Cette bande littorale est de largeur variable, s'étendant jusqu'à 500 mètres du rivage à marée basse. La zone comprend principalement des plages surélevées, en grande partie recouvertes de galets de taille moyenne, de structures basaltiques et de moraines latérales et terminales. Le littoral est très irrégulier et se compose d'une série de petites baies formées parmi des caps rocheux.
Cette topographie constitue une limite naturelle pour l'installation des colonies de mammifères marins et de manchots en phase de reproduction, ce qui justifie l'expansion de la ZSPA.
6 (ii) Caractéristiques du milieu naturel
La zone renferme d'importantes valeurs scientifiques du fait de la présence de colonies d'éléphants de mer (Mirounga leonina) en phase de reproduction, de colonies d'otaries de Kerguelen (Arctocephalus gazella) ne se reproduisant pas, et occasionnellement de phoques de Weddell (Leptonychotes weddelli), de phoques crabiers (Lobodon carcinophagus) et de léopards des mers (Hydrurga leptonyx). Environ 400 éléphants de mer du sud femelles et environ 60 mâles adultes se rendent dans la zone avec leur progéniture lors de la saison de reproduction, (Carlini et al. 2006, Negrete 2011), tandis que pendant la saison de mue, 200 à 800 individus de cette espèce arrivent sur la côte de la ZSPA 132. Les groupes d'otaries de Kerguelen ne se reproduisant pas peuvent atteindre un effectif de 300 individus, bien que ce nombre puisse varier considérablement d'une année à l'autre et puisse excéder 1000 (Durante et al 2007).
La zone abrite également d'importantes colonies de manchots papous (Pygoscelis papua) et de manchots Adélie (Pygoscelis adeliae), comprenant respectivement 3 800 et 3 000 couples. La population de pétrels (essentiellement Oceanites oceanicus et, dans une moindre mesure, Fregetta Tropica) atteint quelque 200 couples. Le goéland dominicain (Larus dominicanus), le chionis blanc (Chionis alba), la sterne subantarctique (Sterna vittata), le pétrel géant (Macronectes giganteus) et les labbes (Catharacta sp.) se reproduisent également dans la zone. Étant donné que certains sites de nidification autour de la péninsule Potter changent de position au fil du temps, le calcul des populations n'est qu'une estimation.
Les manchots Adélie et papous sont répartis autour de la pointe Stranger, entre l'abri Elefante et la Saillie 7. Les concentrations de mammifères sont réparties le long de la côte, entre la Saillie 1 et la Saillie 7, et les sites de nidification des pétrels géants sont généralement répartis autour de la colline des Trois frères (située en dehors de la ZSPA) et entre la Saillie 7 et la Saillie 4 (voir Carte 1). On observe dans la zone un développement relativement abondant de communautés végétales dominées par des lichens et des mousses, respectivement sur les versants rocheux et les surfaces plates de paléo-plages.
Caractéristiques naturelles de la flore
La répartition de la végétation correspond à une combinaison de variables liées entre elles : type de substrat, exposition, stabilité des versants et drainage (disponibilité en eau). La péninsule Potter comprend une zone de plusieurs kilomètres carrés libre de neige et de banquise côtière. Les collines des Trois frères sont environnées d'un substrat relativement stable. Les moraines proches du glacier ne sont couvertes que de plantes éparses mais la couverture végétale et la richesse des espèces augmentent en s'éloignant des moraines. Un plateau situé au sud-ouest des collines des Trois frères est recouvert d'une végétation d'une richesse exceptionnelle. Il comporte deux couches de plantes dont la couverture peut atteindre 100 %. Plusieurs espèces de mousses et de lichens observées sur la péninsule Potter se limitent à cette zone. Il est possible d'observer les deux plantes vasculaires indigènes antarctiques, Colobanthus quifensis et Deschampsia antarctica, dans cette zone (Dopchiz et al. 2017A, 2017b) non loin de la côte ou dans des lieux à fortes concentrations de nutriments.
Les mousses pleurocarpes, telles que Sanionia uncinata et Calliergon sarmentosum, sont dominantes, tandis que les rochers sont généralement couverts de lichens croûteux Lecidea sciatrapha. Plus haut sur le versant, où le sol est mieux drainé et la durée d'enneigement plus courte, des mousses formant des coussins, telles que Andreaea regularis et Andreaea gainii, dominent généralement aux côtés de Himantormia lugubris. Des associations de lichens bryophiles, tels que Psoroma hypnorum, et d'un certain nombre de mousses acrocarpes sont également souvent observées. Dans les lieux où l'enneigement dépasse rarement 10 cm, même en hiver, une voûte composée d'une double couche de lichens et de mousses se forme.
La couche supérieure est irrégulière et comporte des lichens frutescents, tels que Usnea aurantiaco-atra, U. antarctica et Pseudephebe pubescens. La couche inférieure se compose d'un groupement de différentes espèces de mousses et de plantes épilithiques. U. aurantiaco-atra et Himantormia lugubris constituent souvent des tapis entrecroisés. (Bubach et al., 2016, Rivera et al. 2018). Des mousses dicranoïdes comme le Chorisodontium aciphyllum et les lichens frutescents formant des coussins, tels que Sphaerophorus globosus, sont présents dans les creux. Le lichen bryophile le plus abondant est Ochrolechia frigida. (Wiencke et al. 1998)
Accès à la zone
Sauf dans le cas d'exceptions pour lesquelles une autorisation a été octroyée, l'accès à la zone doit se faire à pied, depuis la pointe nord, proche de l'héliport de la base Carlini (62°14'17”S, 58º40'42”O), ou depuis l'arrière du versant nord de la colline des Trois frères (voir Carte 1). L'accès à la zone par la mer sur les plages doit être évité lorsque la faune est présente, particulièrement entre octobre et décembre, car cette période coïncide avec les pics d'activité pour l'allaitement des éléphants de mer et la ponte des œufs.
Des informations complémentaires sont disponibles à la section 7 (ii).
6 (iii) Structures à l'intérieur et à proximité de la zone
Structures dans la zone
Refuges : l'abri argentin Elefante se trouve à environ 150 mètres de la côte, à 1 000 mètres au nord-ouest de la pointe Stranger. Il est utilisé par les groupes de recherche qui mènent des activités dans la ZSPA, de mars à octobre. L'abri peut loger 6 personnes maximum (voir la section 7 (ix) sur l'élimination des déchets).
Panneaux : des panneaux signalant l'entrée de la zone protégée sont situés à la pointe Mirounga (proche de l'héliport), à la base nord sur la colline des Trois frères, et dans la zone de plages proche de la Saillie I. Les panneaux affichent des informations concernant l'existence de la ZSPA et l'obligation de détenir un permis d'accès à la zone.
Structures à proximité de la zone
La base Carlini est une station argentine permanente (62°14'S, 58º39'O) située dans l'anse Potter, péninsule Potter, dans la partie sud-ouest de l'île du Roi-George (Isla 25 de Mayo). Elle comporte plusieurs installations, notamment le laboratoire argentin-allemand Dallmann, une initiative entrepreneuriale entre l'Institut Alfred Wegener (AWI) et l'Institut antarctique argentin (IAA).
L'Albatros est un refuge argentin (62°15'09”S, 58°39'23”O / - 62,2525, - 58,65639) situé dans l'anse Potter, péninsule Potter.
Les autres stations proches sont la station coréenne King Sejong (62°13'394”S / 58°47'190”O) et la station polonaise Arctowsky (62°9'586”S, / 58°28'399”O).
6 (iv) Emplacement d'autres zones protégées à proximité
La ZSPA n° 125, péninsule Fildes, île du Roi-George (Isla 25 de Mayo), îles Shetland du Sud, se trouve à environ 20 km vers l'est.
La ZSPA n° 128, rive occidentale de la baie de l'Amirauté, île du Roi-George (Isla 25 de Mayo), îles Shetland du Sud, se trouve à environ 10 km vers le nord-est.
La ZSPA n° 171, pointe Nar?bski (côte sud-est de la péninsule de Barton, île du Roi-George [Isla 25 de Mayo]).
La ZSPA n° 133, pointe Harmonie, île Nelson, se trouve à environ 30 km en direction ouest/sud- ouest.
6 (v) Zones spéciales à l'intérieur de la zone
Il n'y a aucune aire spéciale à l'intérieur de la zone.
7. Critères de délivrance des permis
7 (i) Conditions générales pour l'obtention d'un permis
L'accès à la zone est interdit sauf avec un permis délivré par les autorités nationales compétentes. Les critères de délivrance d'un permis pour entrer dans la zone sont les suivants :
Les activités sont réalisées à des fins scientifiques, de gestion de la ZSPA ou de vulgarisation, qui ne peuvent être menées ailleurs et qui sont en accord avec les objectifs du plan de gestion. Toutes les activités de gestion (inspection, entretien ou révision) sont conformes au plan de gestion. Le personnel autorisé à entrer dans la zone doit être en possession du permis.
Un rapport de visite doit être soumis à l'autorité nationale compétente mentionnée dans le permis au terme de l'activité, conformément aux conditions établies par les autorités nationales qui ont délivré le permis.
Le tourisme et toute autre activité récréative sont interdits dans la zone.
7 (ii) Accès à la zone et déplacements à l'intérieur de celle-ci
Dans la mesure du possible, les déplacements à l'intérieur de la zone doivent s'effectuer à pied, le long des pistes existantes connues du personnel familiarisé avec la zone et de ses visiteurs habitués. Il s'agit de la zone des plages et de la limite supérieure de la zone, au nord-est de la colline des Trois frères.
Les véhicules, quels qu'ils soient, sont interdits à l'intérieur de la zone, sauf lorsqu'il s'agit de véhicules indispensables pour l'entretien du refuge, qui sont pilotés uniquement par le personnel chargé de la logistique et en conformité avec un permis d'accès. Dans ce cas, l'accès à la ZSPA se fait par une pente douce proche du refuge Albatros et les véhicules doivent éviter les zones de végétation ainsi que les concentrations d'oiseaux et de mammifères (voir Carte 1).
L'exploitation d'aéronefs au-dessus de la ZSPA doit suivre, comme norme minimale, les dispositions incluses dans la résolution 2 (2004), " Directives pour l'exploitation d'aéronefs à proximité de concentrations d'oiseaux dans l'Antarctique ". En règle générale, aucun aéronef n'est autorisé à survoler la ZSPA en dessous de 610 mètres (2 000 pieds). Un espacement horizontal de 460 mètres (1/4 mille marin) de la côte doit être maintenu dans la mesure du possible. Les atterrissages d'aéronefs dans la zone sont interdits, sauf en cas d'urgence ou de sécurité aérienne.
L'exploitation de RPA est interdite dans les limites de la zone, sauf si elle a fait l'objet d'un examen préalable, au cas par cas, lors de la procédure d'évaluation d'impact sur l'environnement. Ils ne pourront être utilisés qu'en vertu d'un permis et selon les conditions stipulées dans le permis. Les directives applicables du Traité sur l'Antarctique seront dûment prises en compte lors du processus d'analyse et d'autorisation.
7 (iii) Activités pouvant être conduites à l'intérieur de la zone
Travaux de recherche scientifique qui ne peuvent être menés ailleurs et qui ne porteront pas atteinte à l'écosystème de la zone.
Activités de gestion essentielles, y compris les visites d'évaluation de l'efficacité du plan de gestion et des activités de gestion.
Activités à des fins pédagogiques et de vulgarisation, qui contribuent à la sensibilisation aux activités scientifiques, dans le cadre des Programmes antarctiques nationaux.
Entretien de l'abri Elefante, sauf entre octobre et décembre. Pendant cette période, les activités d'entretien de l'abri doivent être évitées ou, si nécessaire, réduites dans toute la mesure possible et les tâches doivent toujours être réalisées en conformité avec un permis. Cette période est considérée comme particulièrement sensible car elle coïncide avec les pics d'activité pour l'allaitement des éléphants de mer et la ponte des œufs.
7 (iv) Installation, modification ou enlèvement de structures
Aucune nouvelle structure ne doit être construite à l'intérieur de la zone et aucun matériel scientifique ne doit y être installé, sauf à des fins scientifiques ou de gestion indispensables, et sous réserve de la détention d'un permis approprié.
Tout le matériel scientifique installé dans la zone ainsi que toutes les bornes de recherche doivent être approuvés par un permis et clairement étiquetés, indiquant le pays, le nom du chercheur principal et l'année d'installation. Tous ces équipements doivent être de nature telle que le risque de contamination dans la zone, de dommage causé à la végétation ou de perturbation de la faune soit minimal.
Les structures et les installations doivent être enlevées lorsqu'elles ne sont plus nécessaires, ou une fois que le permis est arrivé à expiration, selon le scénario qui se produit en premier lieu. Aucune trace des activités de recherche ne doit subsister une fois que le permis est arrivé à expiration. Si un projet spécifique ne peut être achevé dans les limites du calendrier spécifié par le permis, ces circonstances doivent être stipulées dans le rapport de visite, et une prorogation de la validité du permis autorisant le matériel à rester dans la zone doit être demandée.
Les tentes sont autorisées uniquement pour entreposer des instruments ou du matériel scientifiques, ou pour être utilisées en tant que postes d'observation.
7 (v) Emplacement des camps
Afin d'éviter les perturbations importantes de la faune, et compte tenu du fait qu'il existe des lieux alternatifs de logement, il n'est pas permis de camper dans la ZSPA n° 132. Les porteurs de projets dont la réalisation est autorisée dans la ZSPA peuvent demander à être logés à la base Carlini, en fonction des disponibilités. Si nécessaire à des fins scientifiques, l'abri Elefante (situé dans la zone) ou l'abri Albatros (hors de la zone, mais très proche) peuvent être utilisé. L'utilisation de l'abri Elefante à des fins scientifiques par le personnel autre que celui du Programme antarctique argentin doit être arrangée à l'avance avec ledit programme.
L'emplacement des campements à proximité de la ZSPA est la responsabilité du Programme antarctique national correspondant, mais pour des raisons de sécurité, il est recommandé d'en informer le chef de la station Carlini.
7 (vi) Restrictions relatives aux matériaux et organismes pouvant être introduits dans la zone
L'introduction délibérée d'animaux vivants ou de matières végétales dans la ZSPA est interdite.
Il convient à tout moment de veiller à ce que les précautions nécessaires contre l'introduction involontaire d'espèces non indigènes dans la zone soient adoptées. Il est à noter que les espèces non indigènes sont plus fréquemment et efficacement introduites par les humains. Les vêtements (poches, bottines, fermetures velcro sur les habits) et l'équipement personnel (sacs, sacs à dos, sacs d'appareils photos, trépieds), ainsi que les instruments scientifiques et les outils de travail, peuvent porter des larves d'insectes, des semences, des propagules, etc. Pour plus d'informations, il convient de se rapporter au " Manuel sur les espèces non indigènes - CPE 2016 ".
L'introduction de produits à base de volaille non cuits dans la zone est interdite.
Aucun herbicide ni pesticide ne sera introduit dans la zone. Tout autre produit chimique introduit avec l'autorisation d'un permis correspondant doit être enlevé de la zone au plus tard avant la fin des activités prévues par le permis. L'utilisation et le type de produits chimiques doivent être documentés avec un maximum de détails pour que d'autres chercheurs puissent en prendre connaissance.
Il est interdit d'entreposer dans la zone du carburant, des denrées alimentaires et d'autres matériaux, sauf s'ils sont absolument nécessaires pour mener à bien l'activité autorisée dans le permis et à condition de les stocker à l'intérieur ou aux alentours de l'abri Elefante et de les enlever à la fin de l'activité. Tout carburant utilisé à l'abri Elefante doit être manipulé conformément au plan d'urgence établi par le Programme antarctique argentin pour la station Carlini.
7 (vii) Prélèvement de végétaux, capture d'animaux ou perturbations nuisibles de la faune et la flore
Tout prélèvement de végétaux, toute capture d'animaux, ou toute perturbation nuisible à la flore et à la faune indigènes est interdit, sauf si un permis l'autorise.
Les distances d'approche de la faune doivent être respectées, sauf si les projets scientifiques ont d'autres exigences, et sous réserve que les permis adéquats aient été délivrés.
Pour les manchots, la distance recommandée est de 10 mètres pendant les périodes de reproduction et de mue, et de 5 mètres par rapport aux jeunes manchots. Une distance de 100 mètres est recommandée par rapport aux nids de pétrels géants et, dans le cas des otaries de Kerguelen, des phoques de Weddell, des léopards de mer et des phoques crabiers, une distance de 10 mètres minimum doit être respectée. Il convient de noter que ces distances servent d'orientation et doivent être adaptées et augmentées si la proximité humaine s'avère être clairement une source de tension pour l'animal.
Dans le cas d'une activité qui implique des prélèvements ou des perturbations nuisibles, celle-ci doit être réalisée conformément à la dernière version disponible du Code de conduite du SCAR pour l'utilisation d'animaux à des fins scientifiques dans l'Antarctique, comme norme minimale.
Les informations concernant les prélèvements et les perturbations nuisibles sont à échanger, comme il convient, par le biais du système d'échange d'informations du Traité sur l'Antarctique et leurs archives sont à déposer, comme norme minimale, auprès du Répertoire maître de l'Antarctique ou en Argentine, auprès du Centre national de données antarctiques (Centro de Datos Nacionales Antárticos).
Les scientifiques qui prélèvent des échantillons de tout type doivent d'adresser au SEEI (Système électronique d'échange d'informations) et/ou contacter les Programmes antarctiques nationaux concernés afin de minimiser le risque d'une éventuelle répétition.
7 (viii) Collecte ou retrait de matériaux non introduits dans la zone par le titulaire du permis
Le ramassage ou l'élimination de matériaux présents dans la zone ne peut être autorisé qu'avec le permis adéquat. L'enlèvement de spécimens biologiques morts à des fins scientifiques doit être examiné au cas par cas, afin de ne pas dépasser des niveaux qui entraîneraient la dégradation de la base nutritive des détritivores locaux. Cette action dépend de l'espèce à ramasser et, si nécessaire, les conseils d'un spécialiste sont requis avant la délivrance du permis.
7 (ix) Elimination des déchets
Tous les déchets non physiologiques doivent être retirés de la zone. Les eaux usées et les résidus liquides domestiques peuvent être déversés en mer, conformément à l'article 5 de l'annexe III du Protocole de Madrid.
Les déchets qui résultent des activités de recherche menées dans la zone peuvent être entreposés à titre temporaire à côté de l'abri Elefante en attendant d'être évacués, à condition qu'ils ne se répandent pas et que la faune n'y ait pas accès. Ces déchets seront acheminés le plus souvent possible à la base Carlini ou éliminés par le programme antarctique concerné en conformité avec l'annexe III du Protocole de Madrid.
7 (x) Mesures qui peuvent être nécessaires pour continuer de répondre aux objectifs du plan de gestion
Des permis d'accès à la zone peuvent être délivrés afin d'y réaliser des activités de biosurveillance et d'inspection du site pouvant impliquer le prélèvement de matières végétales ou d'échantillons d'animaux à des fins scientifiques, ainsi que l'érection ou l'entretien des panneaux et toute autre mesure de gestion.
7 (xi) Rapports de visite
Pour chaque permis délivré, et une fois que les activités sont terminées, le principal détenteur du permis soumet un rapport décrivant les activités menées dans la zone. Ce rapport doit respecter le format convenu et être envoyé ensuite avec le permis à l'autorité qui a délivré le permis.
Les informations comprises dans les rapports seront utilisées dans toute révision du plan de gestion et dans l'organisation de l'usage scientifique de la zone.
Les archives de permis de la ZSPA et les rapports de visite sont échangés avec les autres Parties consultatives, dans le cadre du Système d'échange d'informations, tel que stipulé dans l'article 10.1 de l'annexe V.
Ces rapports doivent être conservés et disponibles en cas d'inspection effectuée par toutes les Parties intéressées, le SCAR, la CCAMLR et le COMNAP, de manière à fournir les informations nécessaires sur les activités humaines menées dans la zone en vue de sa gestion adéquate.
8. Bibliographie
Abele, D., Vazquez, S., Buma, A. G., Hernandez, E., Quiroga, C., Held, C., … & Mac Cormack, W. P. (2017). Pelagic and benthic communities of the Antarctic ecosystem of Potter Cove : Genomics and ecological implications. Marine genomics, 33, 1-11.
Ainley, D.G., Ballard, G., Blight, L.K., Ackley, S., Emslie, S.D., Lescroël, A., Olmastroni, S., Townsend, S.E., Tynan, C.T., Wilson, P., Woehler, E. 2010. Impacts of cetaceans on the structure of southern ocean food webs. Mar. Mam. Sci. 26: 482-489.
Ainley, D.G., Blight, L.K. 2009. Ecological repercussions of historical fish extraction from the Southern Ocean. Fish Fisheries 10: 13-38.
Atkinson, A., Siegel, V., Pakhomov, E., Rothery, P. 2004. Long-term decline in krill stock and increase in salps within the Southern Ocean. Nature 432: 100-103.
Bers, V., Momo, F., Schloss, I.R., Abele, D. (2013) Analysis of trends and sudden changes in environmental long-term data from King George Island (Antarctica) : Relationships between global climatic oscillations and local system response. Climatic Change, en ligne, 11 août 2012. doi : 10.1007/s10584-012-0523-4.
Bubach D, Perez Catán S, Di Fonzo C, Dopchiz L, Arribere M & Ansaldo M. , 2016. Elemental composition of Usnea sp lichen from Potter Peninsula, 25 de Mayo (King George) Island, Antarctica. Environmental Pollution 210 : 238-245. ISSN : 0269-7491).
Carlini A.R., Poljak S., Daneri G.A., Márquez M. E.I., Negrete J. (2006). The dynamics of male harem dominance in southern elephant seals (Mirounga leonina) at the South Shetland Islands. Polar Biology Vol. 29 (10) 796-805.
Carlini A.R., Coria N.R., Santos M. M., Negrete J., Juares M. A., Daneri G.A. 2009. Responses of Pygoscelis adeliae and P. papua populations to environmental changes at Isla 25 de Mayo (King George Island). Polar Biology 32: 1427-1433.
Carlini A.R., Daneri G.A., Márquez M. E.I., Negrete J., Mennucci J., Juares M. 2010. Food consumption estimates of southern elephant seal females at Isla 25 de Mayo (King George Island), Antarctica. XXXIe Conférence du Comité scientifique sur la recherche antarctique et la science ouverte. Buenos Aires, Argentine.
Casaux, R. J., Barrera-Oro, E.R. 2006. Shags in Antarctica : their feeding behaviour and ecological role in the marine food web. Antarctic Science 18: 3-14.
Curtosi, A., Pelletier, E., Vodopivez, C., St Louis, R., Mac Cormack, W. Presence and Distribution of Persistent Toxic Substances in Sediments and Marine Organisms of Potter Cove, Antarctica. Arch Environ Contam Toxicol (2010) 59: 582-592. DOI 10.1007/s00244-010-9509-2.
Daneri G.A., Carlini A.R.1999. Spring and summer predation on fish by Antarctic fur seal, Arctocephalus gazella, at King George Island, South Shetland Islands. Canadian J. of Zoology 77 : 1165-1170.
Di Fonzo C, Zappala C, Cebuhar J y Ansaldo M. , 2014. Stress levels in Pygoscelis papua: a comparison between nesting and molting stages. III APECS-Brazil, September 22 - 26. Libro de Resumos del III APECS Brasil. Pages 56-58. Link : http://www.apecsbrasil.com/news/livro-de-resumos-do-iii-simposio-da-ape…?
Di Fonzo, C. I., Dopchiz, L. P. y M. Ansaldo, 2017a. Bioquímica sanguínea de tres poblaciones antárticas de Pygoselis papua. Guaiquil, I., Leppe, M. , Rojas, P., y R. Canales, Eds. Visiones de Ciencia Antártica, Libro de Resúmenes, IX Congreso Latinoamericano de Ciencias Antártica, Punta Arenas-Chile. Publicación del Instituto Antártico Chileno. Pages 282-285.
Di Fonzo C, Bubach D, Dopchiz L, Arribere M, Ansaldo M, Perez Catan S., 2017b. Plumas de pingüino como bioindicadores de riesgo a elementos tóxicos en ambientes marinos costeros de la isla 25 de Mayo, Antártida. Livre des résumés de la 12e Réunion de la Société de Toxicologie et de Chimie de l'Environnement (SETAC- Amérique latine), page 71.
Dopchiz, L.P., Di Fonzo C.I. y M. Ansaldo, 2017a. Densidad e índice de estomas en Deschampsia antarctica expuesta a impacto antrópico. Guaiquil, I., Leppe, M. , Rojas, P., y R. Canales, Eds. Visiones de Ciencia Antártica, Libro de Resúmenes, IX Congreso Latinoamericano de Ciencias Antártica, Punta Arenas-Chile. Publicación del Instituto Antártico Chileno. Pages 294-296.
Dopchiz LP, Di Fonzo CI, Ansaldo M. , 2017b. Mitotic activity biomarkers in Deschampsia antarctica from different polluted and unpolluted sites. Livre des résumés de la 12e Réunion de la Société de Toxicologie et de Chimie de l'Environnement (SETAC- Amérique latine), page 28.
Durante Martín R., Rossi J.A, Ciai D.N. Daneri G., Pfoh M. 1, y Javier Negrete. Abundancia de lobo fino antártico (Arctocephalus gazella) durante la época post reproductiva en la isla 25 de Mayo, Islas Shetland del Sur, Antártida. VII Jornadas de Jóvenes Investigadores y Extensionistas, 30 de Agosto y 1 de Septiembre de 2017, La Plata, Argentina.
Ducklow, H. W., Baker, K., Martinson, D.G., Quetin, L.B., Ross, R.M., Smith, R.C., Stammerjohn, S.E., Vernet, M. , Fraser. W. 2007. Marine pelagic ecosystems : the West Antarctic Peninsula. Phil. Trans. Roy. Soc. Lond. Ser. B 362: 67-94.
Lignes directrices sur l'exploitation d'aéronefs. résolution 2. - XXVIIe RCTA - VIIe CPE, Le Cap (disponible sur : http://www.ats.aq/documents/recatt/Att224_f.pdf?))
Fuentes, V., Alurralde, G., Meyer, B. Aguirre, G., Canepa, A., Wölfl, A.C., Hass, H.C., Williams, G.N. and Schloss, I.R. (2016) Glacial melting : an overlooked threat to Antarctic krill. Rapports scientifiques 6, 27234 ; doi : 10.1038/srep27234 (2016).
Garcia, M. D., Hoffmeyer, M. S., López Abbate, M. C., Barría de Cao, M. S., Pettigrosso, R.E., Almandoz, G.O., Hernando, M. P., Schloss, I.R. (2015) Micro- and mesozooplankton responses during two contrasting summers in coastal Antarctic environment. Polar Biology. DOI 10.1007/s00300-015-1678-z.
Hernando, M. P., Schloss, I.R., Malanga, G.F., Almandoz, G.O., Ferreyra, G.A., Aguiar, M. B., Puntarulo, S. (2015) Effects of salinity changes on coastal Antarctic phytoplankton physiology and assemblage composition. Journal of Experimental Marine Biology and Ecology, 466 : 110-119.
Marschoff, E.R., Barrera-Oro, E.R., Alescio, N.S., Ainley, D. G. 2012. Slow recovery of previously depleted demersal fish at the South Shetland Islands, 1983-2010. Fisheries Research., 125-126, pp : : 206-213.
Montes-Hugo, M. , Doney, S.C., Ducklow, H.W., Fraser, W., Martinson, D., Stammerjohn, S.E., Schofield,
O. 2009. Recent changes in phytoplankton communities associated with rapid regional climate change along the western Antarctic Peninsula. Science 323: 1470-1473.
Morgan, F., Barker, G., Briggs, C., Price, R. and Keys, H. 2007. Rapport final sur les domaines environnementaux d'Antarctiqe version 2.0, Manaaki Whenua Landcare Research New Zealand Ltd, pp. 89.
Negrete Javier (2011) Estructura, dinámica, mediaciones y consecuencias de las interacciones agonísticas entre machos de elefante marino del sur (Mirounga leonina) en la isla 25 de Mayo, Antártida. 201 pp. Tesis Doctoral. PREBI-SEDICI http://hdl.handle.net/10915/5319?
Manuel sur les espèces non indigènes. XXXIVe RCTA - XIVe CPE, Buenos Aires (disponible sur :
http://www.ats.aq/documents/recatt%5Catt483_f.pdf?))
Rombolá, E. F., Marschoff, E., Coria, N. 2010. Inter-annual variability in Chinstrap penguin diet at South Shetland and South Orkneys Islands. Polar biology. 33 (6), 799-806.
Rivera M. S., Perez Catán S., Di Fonzo C., Dopchiz L., Arribere M. A., Ansaldo M. , Messuti M. I. and Bubach
D.F. 2018. Lichenized fungi as biomonitor of atmospheric elemental composition from Potter Peninsula, 25 de Mayo (King George) Island, Antarctica. Atmospheric Pollution Research (Recherche sur la pollution
atmosphérique). Accepté, révisé et en phase de correction.
Russell, J.L., Dixon, K.W., Gnanadesikan, A., Stouffer, R.J., Toggweiler, D.J.R., 2006. The Southern Hemisphere westerlies in a warming world : propping open the door to the deep ocean. J. Clim. 19: 6382-6390. Stammerjohn, S.E., Martinson, D.G., Smith, R.C., Yuan, X., Rind, D., 2008. Trends in Antarctic annual sea ice retreat and advance and their relation to El Niño-Southern Oscillation and Southern Annular Mode variability.
J. Geophys. Res., 113: C03S90.
Sahade, R., Lagger, C., Torre, L., Momo, F., Monien, P., Schloss, I., Barnes, DKA, Servetto, N., Tarantelli, S., Tatián, M. , Zamboni, N., Abele, D. (2015) Climate change and glacier retreat drive shifts in an Antarctic benthic ecosystem. Science Advances 2015 ; 1 : e1500050.
Schloss, I.R., A. Wasilowska, D. Dumont, G.O. Almandoz, M. P. Hernando, C.-A. Michaud-Tremblay, L. Saravia, M. Rzepecki, P. Monien, D. Monien, E.E. Kopczy?ska, V. Bers, G.A. Ferreyra (2014). On the phytoplankton bloom in coastal waters of southern King George Island (Antarctica) in January 2010 : An exceptional feature ? Limnology & Oceanography 59 (1) : 195-210.
Schloss, I.R., Abele, D., Ferreyra, G.A., González, O., Moreau, S., Bers, V., Demers, S. (2012) Response of Potter Cove phytoplankton dynamics to long term climate trends. Journal of Marine Systems, 92 : 53-66.
Strelin, J., Heredia, P., Martini, M. A., Kaplan, M. M., & Kuhn, G. (2014). The age of the first Holocene marine transgression in Potter Cove, Isla 25 de Mayo (King George Island), South Shetland Islands.
Terauds, A., Chown, S., Morgan, F., Peat, H., Watts, D., Keys, H., Convey, P. and Bergstrom, D. 2012. Conservation biogeography of the Antarctic. Diversity and Distributions, 22 May 2012, DOI : 10.1111/j.1472-4642.2012.00925.x.
Thompson, D.W.J., Solomon, S., 2008. Interpretation of recent Southern Hemisphere climate change. Science 296: 895-899.
Torre, L., Servetto, N., Eöry, L. M. , Momo, F., Abele, D., Sahade, R. 2012.Respiratory responses of three Antarctic ascidians and a sea pen to increased sediment concentrations. Polar biology 35(11) : 1743-1748.
Trivelpiece, W.Z., Hinke, J.T. Miller, A.K. Reiss, C.S. Trivelpiece, S.G., Watters, G.M., 2010. Variability in krill biomass links harvesting and climate warming to penguin population changes in Antarctica. Proc. Natl. Acad. Sci., doi/10.1073/pnas.1016560108.
Vodopivez, C., Curtosi, A., Villaamil, E., Smichowski, P., Pelletier, E., Mac Cormack, W. Heavy metals in sediments and soft tissues of the Antarctic clam Laternula elliptica : More evidence as a ? possible biomonitor of coastal marine pollution at high latitudes ? . Science of the Total Environment 502 (2015) 375- 384. http://dx.doi.org/10.1016/j.scitotenv.2014.09.031?
Wiencke, C., Ferreyra, C., Arntz, W. and Rinaldi, C. 1998. The Potter Cove coastal ecosystem, Antarctica. Synopsis of research performed within the frame of the Argentinean - German Cooperation at the Dallmann Laboratory and Jubany Station (King George Island, Antarctica, 1991 -1 997). Ber. Polarforsch, 299, pp : 342.
Carte 1. - Zone spécialement protégée de l'Antarctique n° 132, Péninsule Potter.
Annexe
MESURE 4 (2018)
ZONE SPÉCIALEMENT PROTÉGÉE DE L'ANTARCTIQUE N° 147 (VALLÉE ABLATION ET MONT GANYMÈDE, ÎLE ALEXANDRE) : PLAN DE GESTION RÉVISÉ (ENSEMBLE UNE ANNEXE)
Les Représentants,
Rappelant les articles 3, 5 et 6 de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement, qui disposent de la désignation des zones spécialement protégées de l'Antarctique (« ZSPA ») et de l'adoption des plans de gestion pour ces zones ;
Rappelant
- la recommandation XV-6 (1989), qui a désigné la vallée Ablation et le mont Ganymède, île Alexandre, comme site présentant un intérêt scientifique particulier (« SISP ») n° 29 et a annexé le plan de gestion du site ;
- la résolution 3 (1996), qui a prorogé la date d'expiration du SISP n° 29 ;
- la mesure 2 (2000) qui a prorogé la date d'expiration du plan de gestion du SISP nº 29 ;
- la décision 1 (2002), qui a renommé et renuméroté le SISP n° 29 en ZSPA n° 147 ;
- la mesure 1 (2002) et la mesure 10 (2013), qui ont adopté des plans de gestion révisés pour la ZSPA n° 147 ;
Rappelant que la recommandation XV-6 (1989) et la résolution 3 (1996) ont été déclarées caduques par la mesure 1 (2011) ;
Rappelant que la mesure 2 (2000) n'est pas entrée en vigueur et a été retirée par la mesure 5 (2009) ;
Notant que le Comité pour la protection de l'environnement a approuvé un plan de gestion révisé pour la ZSPA n° 147 ;
Souhaitant remplacer le plan de gestion actuel pour la ZSPA n° 147 par le plan de gestion révisé ;
Recommandent à leurs Gouvernements d'approuver la mesure suivante conformément au paragraphe 1 de l'article 6 de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement :
Que :
1. Le plan de gestion révisé pour la zone spécialement protégée de l'Antarctique n° 147 (vallée Ablation et mont Ganymède, île Alexandre), qui figure en annexe de cette mesure, soit approuvé ; et
2. Le Plan de gestion pour la zone spécialement protégée de l'Antarctique n° 147 annexé à la mesure 10 (2013) soit abrogé.
Annexe
PLAN DE GESTION POUR LA ZONE SPÉCIALEMENT PROTÉGÉE DE L'ANTARCTIQUE N° 147
VALLÉE ABLATION ET MONT GANYMÈDE, ÎLE ALEXANDRE
Introduction
La désignation de la vallée Ablation et mont Ganymède, île Alexandre (70°48' S, 68°30' O, environ 180 km2) comme zone spécialement protégée de l'Antarctique (ZSPA) est motivée principalement par la protection des valeurs scientifiques, notamment géologiques, géomorphologiques, glaciologiques, limnologiques et écologiques de cette vaste zone d'ablation.
La vallée Ablation et le mont Ganymède, île Alexandre ont été au départ désignés en 1989 site d'intérêt scientifique particulier (SISP) n° 29 Pointe Ablation - mont Ganymède, île Alexandre en vertu de la recommandation XV-6, proposée par le Royaume-Uni. Ce site portait sur une région libre de glace dans sa majeure partie et située, d'une part, entre les latitudes 70°45' S et 70°55' S et, d'autre part, la longitude 68°40' O et la côte du goulet George VI. La zone comprend plusieurs systèmes de vallées séparés par des crêtes et un plateau d'une altitude comprise entre 650 et 760 mètres. Le plan de gestion initial (recommandation XV-6) décrivait la zone comme « une des zones de plus forte ablation en Antarctique occidental… [avec] … une géologie complexe, les principaux types de roches étant des conglomérats, des schistes et des grès arkosiques, comprenant notamment des schistes boueux et graveleux ainsi que des brèches sédimentaires. La base de la succession est formée d'un mélange spectaculaire qui comprend de grands amas de lave et d'agglomérats. Celle-ci affleure sur les lits de la vallée ainsi qu'au pied de plusieurs falaises. [La zone] possède de nombreuses particularités géomorphologiques, notamment des plages surélevées, des systèmes de moraines et des sols réticulés. Elle renferme également plusieurs lacs d'eau douce gelés en permanence et plusieurs lagunes qui, elles, sont libres de glace et abritent une flore (y compris des bryophytes aquatiques) et une faune très variées. La végétation est en général peu abondante, le seul type de communauté dominée par la mousse et l'hépatique étant limité aux « oasis » où on décèle la présence d'eau dans un cadre montagneux en général sec et désolé. Les écosystèmes d'eau douce et terrestres sont vulnérables à l'action de l'homme et, par conséquent, méritent d'être protégés de toute présence humaine excessive ». En résumé, les principales valeurs de la zone sont ses particularités écologiques, limnologiques, glaciologiques, géomorphologiques et géologiques ainsi que son remarquable intérêt scientifique connexe puisqu'il s'agit d'une des plus grandes zones d'ablation sans glace de l'Antarctique occidental. La zone a été renumérotée ZSPA n° 147 par le biais de la décision 1 (2002) et un plan de gestion révisé a été adopté par le biais de la mesure 1 (2002).
La ZSPA n° 147 vallée Ablation et mont Ganymède, île Alexandre s'intègre dans le contexte plus large du système de zones protégées de l'Antarctique en protégeant l'une des plus grandes zones d'ablation en Antarctique occidental. Des valeurs environnementales et scientifiques équivalentes ne sont pas protégées dans d'autres ZSPA dans la région de la péninsule antarctique. La résolution 3 (2008) recommandait que l'Analyse des domaines environnementaux pour le continent antarctique serve de modèle dynamique pour l'identification des zones spécialement protégées de l'Antarctique dans le cadre environnemental et géographique systématisé visé à l'article 3 (2) de l'annexe V du Protocole (voir également Morgan et al., 2007). Si l'on s'en tient à ce modèle, de petites parties de la ZSPA 147 se trouvent dans le domaine environnemental E (péninsule antarctique et principaux champs de glace de l'île Alexandre) ; pourtant, bien que cela ne soit pas mentionné de manière spécifique dans Morgan et al. il se peut que la zone englobe le domaine C (Géologique du sud de la péninsule antarctique). D'autres zones protégées englobant le domaine E comprennent les ZSPA nos 113, 114, 117, 126, 128, 129, 133, 134, 139, 149, 152, 170, ainsi que les ZSGA 1 et 4. D'autres zones protégées contenant le domaine C comprennent la ZSPA 170 (même si Morgan et al., 2007 ne l'a pas relevé de manière spécifique). La ZSPA fait partie de la Région de conservation biogéographique de l'Antarctique (RCBA) 4, sud central de la péninsule antarctique, et constitue l'une des deux ZSPA de la RCBA 4, l'autre étant la ZSPA N° 170 (Terauds et al., 2012 ; Terauds et Lee, 2016).
1. Description des valeurs à protéger
Les valeurs faisant partie du plan de gestion initial sont réaffirmées dans le présent plan de gestion. D'autres valeurs qui ressortent avec évidence de descriptions scientifiques de la vallée Ablation et mont Ganymède sont autant de raisons importantes justifiant une protection spéciale de la zone. Ces valeurs sont :
La présence d'affleurements sur la formation Fossil Bluff, structure géologique d'importance capitale, car c'est la seule région connue présentant un affleurement ininterrompu englobant le Jurassique et le Crétacé dans l'Antarctique et, dès lors, un lieu essentiel pour comprendre les changements qu'ont connus la flore et la faune entre ces deux ères géologiques.
La présence d'un phénomène géomorphologique contigu, au caractère unique et exceptionnel, révélant des fluctuations de glaciers et de la plateforme glaciaire sur plusieurs milliers d'années, ainsi que d'autres traits géomorphologiques issus de processus glaciaires, périglaciaires, lacustres, éoliens, alluviaux et érosifs.
Deux lacs d'eau douce gelés en permanence (lacs Ablation et Moutonnée) qui ont la particularité d'être en contact avec les eaux salées du goulet George VI.
La présence d'un biote marin, y compris de poissons Trematomus bernacchii, dans le lac Ablation où plusieurs phoques ont été observés malgré la distance de près de 100 km qui les sépare de la haute mer.
La zone recèle une diversité de bryophytes (au moins 21 espèces) qui n'existe sur aucun autre site sous cette latitude en Antarctique ; elle recèle aussi un biote de cyanobactéries, d'algues et de lichens (plus de 35 taxons). La plupart des bryophytes et des lichens se trouvent surtout à la limite méridionale de leur aire de répartition. Certaines de ces espèces sont extrêmement rares en Antarctique.
Plusieurs mousses sont observées dans les lacs et les lagunes à des profondeurs de 9 m. Bien que ces espèces soient toutes terrestres, elles peuvent vivre submergées pendant plusieurs mois chaque année lorsque leur habitat est inondé. Une espèce, Campylium polygamum, s'est adaptée à la vie aquatique et certaines colonies, en immersion permanente, ont atteint des dimensions considérables pouvant dépasser 30 cm. Elles représentent le plus bel exemple de végétation aquatique dans la région de la péninsule antarctique.
Plusieurs espèces de bryophytes sont fertiles dans la zone (produisant des sporophytes) et certaines sont d'ailleurs inconnues ou très rares ailleurs en Antarctique dans ce contexte spécifique (hépatique Cephaloziella varians et mousses Bryoerythrophyllum recurvirostrum, Distichium cappilaceum, Schistidium spp.).
La zone abrite les plus importantes concentrations de végétation de l'île Alexandre. Plusieurs se retrouvent notamment dans les zones irriguées où les communautés de bryophytes et de lichens couvrent jusqu'à 100 m2, voire plus. Dans les zones irriguées peu exposées, des groupes d'espèces terricoles développent des communautés inconnues ailleurs en Antarctique, tandis que les crêtes de roches exposées et les champs de galets stables abritent une communauté localement abondante de lichens, en général dominée par l'espèce Usnea sphacelata.
A titre de comparaison, la zone est riche en nombre et en abondance des espèces microarthropodes si l'on considère leur répartition aussi loin au sud. Le collembole Friesia topo y est particulièrement représenté et tout porte à croire qu'il est endémique à l'île Alexandre. La vallée Ablation est le seul site de l'île Alexandre où l'acarien prédateur Rhagidia gerlachei a été observé, rendant le réseau trophique plus complexe qu'à d'autres endroits sous cette latitude.
2. Buts et objectifs
Les buts et objectifs du plan de gestion sont les suivants :
- éviter toute détérioration ou tout risque de détérioration des valeurs de la zone en empêchant toute perturbation humaine inutile de ladite zone ;
- éviter ou réduire au maximum l'introduction de plantes, d'animaux et de microorganismes non indigènes dans la zone ;
- permettre d'effectuer des recherches scientifiques dans la zone, pour autant qu'elles soient indispensables, qu'elles ne puissent être menées ailleurs et qu'elles ne portent pas atteinte à l'écosystème naturel de la zone ; et
- préserver l'écosystème naturel de la zone afin que celle-ci puisse servir de zone de référence dans les études ultérieures.
3. Activités de gestion
Les activités de gestion ci-dessous seront menées à bien afin de protéger les valeurs de la zone :
Les bornes, les panneaux ou autres structures (p. ex. cairns) érigés dans la zone à des fins scientifiques et de gestion seront attachés et maintenus en bon état puis enlevés lorsqu'ils ne sont plus nécessaires.
Des copies de ce plan de gestion doivent être mises à la disposition des aéronefs prévoyant de visiter les abords de la zone.
Le plan de gestion sera réexaminé au moins tous les 5 ans et mis à jour en conséquence. Un exemplaire du présent Plan de gestion sera mis à la disposition de la station de recherche de Rothera (Royaume-Uni, 67°34' S ; 68°07' O) et de la station General San Martin (Argentine, 68°08' S ; 67°06' O).
Toutes les activités scientifiques et de gestion entreprises au sein de la zone doivent faire l'objet d'une évaluation d'impact sur l'environnement conformément à ce que requiert l'annexe I du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement.
Les directeurs des programmes antarctiques nationaux en cours d'exécution dans la région se livreront entre eux à des consultations pour veiller à ce que les activités de gestion susmentionnées soient mises en œuvre.
4. Durée de désignation
La zone est désignée pour une période indéterminée.
5. Cartes et photographies
Carte 1. - Coordonnées de la vallée Ablation et mont Ganymède, dans la péninsule antarctique. Spécifications de la carte : WGS84 Stéréographique polaire antarctique. Méridien central : - 55° : Parallèle de référence : - 71°.
Carte 2. - Coordonnées de la ZSPA N° 147, vallée Ablation et mont Ganymède, île Alexandre Spécifications de la carte : WGS 1984 Stéréographique polaire antarctique. Méridien central : - 71° ; Parallèle de référence : - 71°.
Carte 3. - Carte topographique de la ZSPA N° 147, vallée Ablation et mont Ganymède, île Alexandre Spécifications de la carte : WGS 1984 Stéréographique polaire antarctique. Méridien central : -68,4° ; Parallèle de référence : -71,0°.
6. Description de la zone
6 (i) Coordonnées géographiques, bornage et caractéristiques du milieu naturel
Description générale
La vallée Ablation et mont Ganymède (zone délimitée par les latitudes 70°45' S et 70°55' S et les longitudes 68°21'O et 68°40'O, environ 180 km2) est située sur la côte est de l'île Alexandre, la plus grande des îles au large de la côte occidentale de la terre Palmer sur la péninsule antarctique (cartes 1 et 2). La distance ouest-est au centre de la zone est d'environ 10 km tandis que la distance nord-sud est de 18 km environ. La zone est entourée, à l'ouest, par la partie supérieure du glacier Jupiter, à l'est par la plateforme glaciaire du goulet George VI, au nord par le glacier Grotto et, au sud, par les avancées inférieures du glacier Jupiter. La vallée Ablation et le mont Ganymède abritent la plus importante zone libre de glace contiguë du secteur péninsulaire antarctique. Elle compte aussi des champs de glace permanents plus petits et des glaciers encaissés qui occupent à peine 17 % de la superficie de la zone. Caractérisée par une topographie montagneuse, la zone comprend des vallées escarpées séparées par des crêtes légèrement ondulantes aux allures de plateaux qui se situent à une altitude moyenne de 650 à 750 mètres, pouvant atteindre parfois 1 070 mètres (Clapperton et Sugden, 1983). La région a fait l'objet d'une glaciation importante même si la structure relativement plate des roches sédimentaires et les rigoureuses conditions climatiques ont contribué à une topographie plutôt arrondie à l'exception de quelques « marches » de falaises constituées de conglomérats et de grès en couches importantes (Taylor et al., 1979).
La zone comprend quatre vallées principales libres de glace (Ablation, Moutonnée, Flatiron et Striation) dont les trois premières contiennent de grands lacs d'eau douce couverts de glace (Heywood, 1977, Convey et Smith, 1997). Le plus grand d'entre eux est le lac proglaciaire Ablation (environ 7 km2) qui a été retenu par les glaces « remontant » la vallée sous la pression du déplacement vers l'ouest de l'épaisse plateforme glaciaire George VI de 100 à 500 mètres d'épaisseur dont la surface se situe à 30 m au-dessus du niveau de la mer (Heywood, 1977, Clapperton et Sugden, 1982). D'un point de vue biologique, l'écosystème terrestre est à un point intermédiaire entre l'Antarctique maritime plutôt doux du nord et l'Antarctique continental plus sec et plus froid de la partie méridionale. En dépit de sa « sécheresse », la vallée abrite un biote extrêmement riche et constitue un outil de référence très utile pour les comparaisons avec les zones d'ablation plus extrêmes et biologiquement plus pauvres du continent Antarctique (Smith, 1988).
Limites
La zone désignée comprend la totalité du massif abritant la vallée Ablation et mont Ganymède qui est délimité à l'ouest par la crête principale séparant le glacier Jupiter des principales vallées Ablation-Montonnée-Flatiron (carte 3). La limite orientale se situe à la bordure orientale de la plateforme glaciaire George VI. La limite septentrionale de la zone est définie par la crête principale séparant le glacier Grotto de la vallée Erratique et d'autres vallées tributaires de la vallée Ablation directement au sud. Au nord-ouest de la zone, la limite longe le col - majoritairement recouvert de glaces - qui sépare la partie supérieure du glacier Jupiter de la vallée Ablation. La limite méridionale de la zone, allant de l'est de la crête principale sur le flanc occidental de la vallée Flatiron jusqu'au point de jonction entre le glacier Jupiter et la plateforme glaciaire George VI, correspond au bord septentrional du glacier Jupiter. Comme, à certains endroits, la ligne de démarcation entre le lac Ablation et la plateforme glaciaire George VI est floue, la limite orientale de la zone au niveau de la vallée Ablation se définit comme une ligne droite orientée plein sud entre l'extrémité est de pointe Ablation et le point de contact entre la plateforme glaciaire et la terre, point à partir duquel la limite orientale suit la lisière terre-glacier. La physiographie est semblable plus au sud, au lac Moutonnée, et la limite orientale à cet endroit se définit comme une ligne droite s'étendant de l'extrémité est de la pointe sur le flanc nord du lac Moutonnée (et l'entourant partiellement) à un endroit abritant un grand bassin d'eau de fonte et où la plateforme glaciaire rejoint la terre à partir d'où la frontière suit la lisière terre-glacier du manteau continental vers le sud où se rejoignent le glacier Jupiter et la plateforme glaciaire George VI. Ainsi, la zone comprend la totalité des lacs Ablation et Moutonnée ainsi que les parties de la plateforme glaciaire qui les retiennent. Les coordonnées des limites figurent à l'annexe 1.
Climat
Aucune donnée météorologique approfondie n'est disponible sur la zone de la vallée Ablation et mont Ganymède, mais son climat a été décrit comme étant dominé par la double influence de dépressions cycloniques partant de l'océan Austral en direction de l'est, contre le courant d'air anticyclonique frais, plus continental et allant du nord au nord-ouest, qui part de la plateforme glaciaire de l'ouest de l'Antarctique (Clapperton et Sugden, 1983). Les dépressions cycloniques génèrent un temps relativement doux, des vents violents du nord et un épais couvert nuageux sur la région, tandis que les mouvements anticycloniques produisent des conditions visibles, fraîches et stables, avec des températures au-dessous de 0 °C ainsi que des vents relativement faibles en provenance du sud. Les données enregistrées dans les environs (à 25 km), au début des années 1970, révèlent une température estivale moyenne légèrement au-dessous de zéro et une moyenne annuelle d'environ - 9 °C (Heywood, 1977) ; les précipitations s'évaluaient à < 200 mm d'eau équivalent par an, avec peu de neige en été. On rencontre souvent une légère couche de neige post-hivernale, mais la région demeure généralement sans neige à partir de la fin de l'été, hormis quelques flocons isolés qui peuvent persister par endroits.
Géologie
La géologie de la vallée Ablation et mont Ganymède est complexe mais reste dominée par des roches sédimentaires bien stratifiées. La caractéristique la plus visible du massif reste un grand anticlinal asymétrique, avec une orientation nord-ouest - sud-est et s'étendant du glacier Grotto jusqu'au glacier Jupiter (Bell, 1975 ; Crame et Howlett, 1988). Des failles chevauchantes sur la partie centrale du massif suggèrent des déplacements verticaux de strates jusqu'à 800 mètres (Crame et Howlett, 1988). Les principales lithologies se composent de conglomérats, de grès arkosiques et d'argiles schisteuses fossilifères, avec des boues rocheuses subordonnées et des brèches sédimentaires (Elliot, 1974 ; Taylor et al., 1979 ; Thomson, 1979). Un ensemble de fossiles a été identifié sur les strates, appartenant au Jurassique supérieur/Crétacé inférieur, dont des bivalves, des brachiopodes, des bélemnites, des ammonites, des dents de requin et des plantes (Taylor et al., 1979 ; Thomson, 1979 ; Crame et Howlett, 1988 ; Howlett, 1989). Plusieurs types de laves interstratifiées ont été observés aux plus bas affleurements de la pointe Ablation (Bell, 1975). La base de cette succession est constituée d'un mélange impressionnant de blocs de lave et d'agglomérés qui affleurent aussi bien sur les sols des vallées qu'à la base de nombreuses falaises (voir Bell, 1975 ; Taylor et al., 1979). La présence d'affleurements sur la formation Fossil Bluff, structure géologique d'importance capitale, car c'est la seule région connue présentant un affleurement ininterrompu englobant le Jurassique et le Crétacé dans l'Antarctique est, dès lors, un lieu essentiel pour comprendre les changements qu'ont connus la flore et la faune entre ces deux ères géologiques.
Géomorphologie et sols
Toute la zone était jadis couverte de glaciers venant de l'intérieur de l'île Alexandre. Ainsi, les formes terrestres nées, tant de l'érosion glaciaire que de dépôts, sont présentes partout dans la zone, ce qui prouve l'avènement d'un ancien mouvement général de glace vers l'est dans le détroit George VI (Clapperton et Sugden, 1983). Les glaciers inadaptés, le lit rocheux strié et les blocs erratiques indiquent une considérable déglaciation qui remonte au maximum glaciaire du Pléistocène (Taylor et al., 1979 ; Roberts et al., 2009). De nombreuses moraines terminales présentes sur le front des derniers glaciers actuels, plusieurs sites étonnamment libres d'astragales et les roches moutonnées polies et striées indiquent que le retrait glaciaire a pu être rapide (Taylor et al., 1979). Il est permis de conclure que la plateforme de glace George VI n'existait pas encore entre 9 600 et 7 730 ans environ (année civile, BP), ce qui laisse croire que le massif de la vallée Ablation et mont Ganymède était probablement libre de glaces permanentes à cette époque, même si un certain nombre de mouvements glaciaires ultérieurs ont été enregistrés dans la région (Clapperton et Sugden, 1982 ; Bentley et al., 2005 ; Smith et al., 2007 a, b ; Roberts et al., 2008 ; Bentley et al., 2009). L'absence de plateforme glaciaire laisse penser que les variabilités océaniques-atmosphériques du début de l'Holocène dans la péninsule antarctique étaient bien supérieures à ce qu'on a relevé ces dernières décennies (Bentley et al., 2005). Roberts et al. (2009) ont étudié les deltas adjacents aux lacs Ablation et Moutonnée qui, au départ, étaient plus élevés que le niveau actuel des lacs, et ils en ont conclu que le niveau de la mer avait baissé de quelque 14,4 mètres depuis la mi-Holocène dans cette partie de l'île Alexandre).
Le relief existant au sein de la zone a été modifié par des processus périglaciaires, gravitationnels et fluviaux. Le lit rocheux des surfaces supérieures du plateau (où il a été largement maintenu intact au point d'être surchargé) a été brisé en fragments plats et en blocs sous l'action du gel (Clapperton et Sugden, 1983). Sur les pentes de la vallée, les lobes de gélifluction, blocs et cercles rocheux sont nombreux, alors que les sols de la vallée sont jonchés de cercles rocheux et de formes polygonales sur des sédiments glaciaires et fluvioglaciaires soumis à l'action du gel. Les parois de la vallée sont également dominées par un relief issu de l'action du gel, de l'activité des roches et de la glace, ainsi que des mouvements saisonniers d'eau de fonte, qui ont laissé des pentes d'éboulis omniprésentes et des failles rocheuses visibles sous des ravins incisés. Les énormes débris de roches sédimentaires fissiles ont aussi entraîné le développement (à environ 50°) de fortes pentes rectilignes, horizontales et légèrement couvertes de débris sur le substrat rocheux. Quelques reliefs éoliens ont été relevés, avec des dunes mesurant jusqu'à 1 mètre de haut et de 8 m de long, comme, par exemple, dans la vallée Erratique (Clapperton et Sugden, 1983). De fines couches de tourbe de 10 à 15 mètres de profondeur sont rencontrées parfois dans les zones végétales, traduisant les évolutions les plus notables du sol de la zone.
Ecologie d'eau douce
La vallée Ablation et mont Ganymède est un site limnologique exceptionnel abritant un certain nombre de lacs, d'étangs et de cours d'eau ainsi qu'une flore benthique généralement riche. De fin décembre à février, l'eau ruisselante naît de trois sources, à savoir les précipitations, les glaciers et la fonte de la plateforme glaciaire George VI, et toute cette eau s'écoule généralement vers la côte (Clapperton et Sugden, 1983). La plupart des cours d'eau, d'une longueur de plusieurs kilomètres, recueillent les eaux des glaciers et des champs de neige permanents. Les principaux cours d'eau s'écoulent dans les lacs Ablation et Moutonnée, qui sont tous deux endigués par la plateforme glaciaire. Selon des études menées au début des années 1970, ces lacs gelés avaient une profondeur de 2,0 à 4,5 mètres toute l'année, avec des profondeurs maximales de 117 mètres et 50 mètres respectivement (Heywood, 1977). Dans les deux lacs, une couche supérieure et stable d'eau fraîche, d'environ 60 mètres et 30 mètres respectivement, renferme des eaux de plus en plus salines influencées par l'interconnexion avec l'océan se trouvant sous la plateforme glaciaire, tout en soumettant les lacs à l'influence des marées (Heywood, 1977). Les bassins d'eau de fonte de surface qui, en hiver, forment en particulier des cuvettes entre les chaînes de pression de la glace des lacs, sont en crue quotidiennement et envahissent même les zones alluviales des vallées plus basses (Clapperton et Sugden, 1983).
Des observations récentes indiquent une diminution de la couverture glaciaire permanente des lacs, par exemple de l'ordre de 25 % pour le lac Moutonnée qui n'avait pas de couverture glaciaire durant les étés de 1994-1995 et 1997-1998 (Convey et Smith, 1997 ; Convey comm. perso., 1999). Toutefois, les trois lacs de la zone étaient presque entièrement couverts de glace au début de février 2001 (Harris, 2001). De nombreux étangs et bassins éphémères et généralement allongés se forment sur les côtés de l'écart entre la terre ferme et la plateforme glaciaire et leur longueur varie entre 10 et 1 500 mètres, avec une largeur allant jusqu'à 200 mètres et des profondeurs oscillant entre 1 et 6 mètres (Heywood, 1977 ; Clapperton et Sugden, 1983). Ces étangs/bassins entrent souvent en crue en période de fonte, mais parfois ils peuvent s'écouler subitement à travers des fissures sous-glaciaires vers la plateforme glaciaire et rendant visibles les anciens rivages des lacs sur les moraines environnantes. Les mêmes étangs/bassins peuvent varier énormément en termes de turbidité en fonction de la présence de sédiments glaciaires suspendus. Les bassins sont en général libres de glace en été, tandis que les grands étangs conservent souvent une partie du couvert glaciaire ; il se peut que seule l'eau des étangs profonds se congèle et devienne solide en hiver (Heywood, 1977). De nombreux étangs d'un hectare et de 15 mètres de profondeur sont présents dans les vallées, certains possédant de larges couvertures de mousses mesurant jusqu'à 9 mètres de profondeur (Light et Heywood, 1977). Les espèces dominantes qui y ont été décrites étaient Campylium polygamum et Dicranella, dont les tiges atteignaient 30 cm de longueur. Des espèces telles que le Bryum pseudotriquetrum (et probablement une seconde espèce de Bryum), Distichium capillaceum, ainsi qu'une espèce non identifiée de Dicranella ont toutes évolué sur le substrat benthique à 1 m de profondeur ou plus (Smith, 1988). La couverture en mousses atteignait 40 à 80 % dans la zone de 0,5 à 5 mètres de profondeur (Light et Heywood, 1975). Le gros de la zone restante était couvert de feutres cyanobactériens (11 taxons) ayant jusqu'à 10 cm d'épaisseur, avec une dominance de Calothrix, Nostoc et Phormidium, ainsi que 36 taxons de microalgues dérivées (Smith, 1988). Les augmentations considérables de mousses indiquent que ces étangs sont peut-être relativement permanents, même si leurs niveaux peuvent fluctuer d'une année à l'autre. La température de l'eau atteint quelque 7 °C dans les étangs plus profonds et 15 °C dans les bassins peu profonds en été, offrant aux bryophytes un environnement relativement favorable et stable. Les bassins peu profonds - habitats de plusieurs types de mousses - peuvent normalement abriter une végétation terrestre et être en crue pendant de courtes périodes en été (Smith, 1988). Les algues abondent dans les cours d'eau à faible débit ainsi que dans les eaux de fonte éphémères, même si elles ne colonisent pas les lits instables des cours d'eau à haut débit. Par exemple, les vastes zones humides du niveau du sol, dans la vallée Moutonnée, disposent d'une flore particulièrement riche avec plus de 90 % de couverture à certains endroits, avec cinq espèces de desmidiales (rares en Antarctique) et de nombreux Zygnema filamenteux verts, ainsi que Nostoc spp. et Phormidium spp. dans les zones plus sèches, moins stables et ensablées (Heywood, 1977).
Les protozoaires, les rotifères, les tardigrades et les nématodes constituent la faune benthique des bassins, étangs et cours d'eau (Heywood, 1977). Les densités sont généralement les plus fortes dans les cours d'eau à faible débit. Le copépode Boeckella poppei se trouvait en abondance dans les lacs, étangs et bassins, mais pas dans les cours d'eau. Le poisson marin Trematomus bernacchii était capturé dans des pièges placés dans le lac Ablation à une profondeur de 70 m, dans la couche d'eau saline (Heywood et Light, 1975 ; Heywood, 1977). Une espèce de phoque non identifiée, mais probablement crabier (Lobodon carcinophagus) ou de Weddell (Leptonychotes weddellii) a été signalée à l'extrémité du lac Ablation à la mi-décembre 1996 (Rossaak, 1997), et plusieurs personnes ont déclaré avoir observé des phoques solitaires les saisons précédentes (Clapperton et Sugden, 1982).
Végétation
La zone de la vallée Ablation - mont Ganymède est essentiellement aride. Peu abondante, la végétation est répartie de manière discontinue. Cependant, des communautés végétales complexes existent dans les zones d'infiltration et le long des cours d'eau, et présentent un intérêt tout particulier car :
1. Elles évoluent dans un environnement quasi stérile.
2. Les communautés mixtes de bryophytes et de lichens sont les plus développées et les plus diversifiées au sud de la latitude 70° S (Smith, 1988 ; Convey et Smith, 1997).
3. Certaines unités taxonomiques de bryophytes sont abondamment fertiles et productives, dans leur limite sud, ce qui est un phénomène inhabituel chez toutes les bryophytes de l'Antarctique, notamment à l'extrême sud (Smith et Convey, 2002).
4. La région est la localité méridionale la plus connue de par sa richesse taxonomique ; et
5. Même si certaines de ces communautés sont présentes également à d'autres endroits au sud-est de l'île Alexandre, la zone présente les meilleurs et plus importants exemples connus sous cette latitude.
La diversité de mousses est particulièrement élevée à cette latitude, avec au moins 21 espèces enregistrées dans la zone, soit 73 % des espèces connues dans l'île Alexandre (Smith, 1997). Les lichens sont également présents en grande variété, avec plus de 35 taxons connus. S'agissant des microlichens, 12 des 15 espèces connues dans l'île Alexandre sont représentées dans la zone (Smith, 1997). Les vallées Ablation, Moutonnée et Striation, ainsi que le littoral SE, regorgent de vastes bassins de végétation tant terrestre que marine (Smith, 1997 ; Harris, 2001). Smith (1988, 1997) a rapporté que la végétation de bryophytes se trouve généralement dans des zones de 10 à 50 mètres2, voire 625 m2 pour certaines, et à des altitudes comprises entre environ 5 mètres et 40 mètres, sur des pentes douces situées au nord ou orientées vers l'est, au niveau des vallées principales. Harris (2001) a, pour sa part, enregistré de grands bassins de bryophytes presque contigus pouvant atteindre 8 000 m2, sur des pentes douces orientées sud-est, au niveau du littoral sud-est de la zone, à une altitude de quelque 10 mètres, près du lieu de jonction entre le glacier Jupiter et la plateforme George VI. Un bassin continu d'environ 1 600 m2 a été enregistré sur les pentes humides de la partie inférieure de la vallée Striation. Plusieurs larges bandes continues de mousses (atteignant 1 000 m2) ont été observées sur les pentes de la vallée Striation orientées sud-ouest et nord-ouest, à une hauteur de 300 à 400 mètres. D'autres petites bandes discontinues de mousses ont été enregistrées à proximité, à une altitude de 540 mètres. Des mousses ont été également observées sur les sommets de la vallée Ablation, à des altitudes de 700 mètres environ.
La bryophyte dominante des zones les plus humides est l'hépatique Cephaloziella varians, qui forme un tapis noirâtre de pousses densément entrelacées. Bien que les données recueillies dans la partie la plus méridionale pour l'espèce C. varians aient été enregistrées à 77° S à partir de la baie Botany, à cap Geology (ZSPA n° 154) en terre Victoria, les vastes tapis qu'elle forme dans le massif de la vallée Ablation - mont Ganymède représentent les étendues les plus importantes de cette espèce aussi bien dans l'extrême-sud que dans l'Antarctique maritime. Les cyanobactéries, notamment Nostoc et Phormidium spp., apparaissent fréquemment soit sur une surface occupée par les hépatiques, soit sur le sol ou en compagnie de pousses de mousse. Au-delà des zones les plus humides, des tapis ondulés de mousses pleurocarpes dominées par Campylium polygamum constituent les étendues les plus verdoyantes de la végétation, avec des Hypnum revolutum. Ces tapis occupent jusqu'à 10-15 cm de tourbes constituées en grande partie de pousses moribondes et non décomposées de mousses. Mélangés à ces mousses mais souvent prédominants sur des espaces plus secs, les Bryum pseudotriquetrum poussent comme des coussins isolés qui peuvent fusionner pour former une tourbe alambiquée. Sur ces zones périphériques plus sèches, plusieurs autres bryophytes formant des tourbes se mélangent souvent aux Bryum. Hormis les espèces plus hydriques susmentionnées, celles-ci comprennent le taxon calcicole Bryoerythrophyllum recurvirostrum, Didymodon brachyphyllus, Distichium capillaceum, Encalypta rhaptocarpa, E. procera. Pohlia cruda, Schistidium antarctici, Tortella fragilis. Syntrichia magellanica, Tortella alpicola, ainsi que plusieurs espèces non identifiées de Bryum et de Schistidium.
Le massif de la vallée Ablation - mont Ganymède se distingue par une occurrence inhabituelle d'un certain nombre de bryophytes fertiles. Les bryophytes antarctiques ne produisent guère de sporophytes, mais Bryum pseudotriquetrum, Distichium capillaceum, Encalypta rhaptocarpa, E. procera et Schistidium spp. ont tous été enregistrés comme étant des espèces fréquemment fertiles de la zone. Très rarement, de petites quantités de mousses Bryoerythrophyllum recurvirostre et d'hépatiques Cephaloziella varians ont été observées comme étant productives dans la vallée Ablation et c'était la première fois qu'on le constatait quelque part en Antarctique (Smith comm. perso., cité dans Convey, 1995 ; Smith, 1997 ; Smith et Convey, 2002). En outre, D. capillaceum n'a jamais été enregistré avant avec des sporophytes à travers l'Antarctique maritime (Smith, 1988). E. procera n'a été rapporté comme étant fertile que dans une seule zone de l'Antarctique (sur l'île Signy, îles Orcades du Sud ; Smith, 1988). Au-delà des zones d'infiltration permanentes, la végétation de bryophytes est extrêmement clairsemée et confinée aux habitats disposant d'eau au moins quelques semaines durant l'été. Ces sites apparaissent de manière sporadique sur les sols de la vallée, avec des rayures de roches sur les pentes, ainsi que dans les fissures qui se forment sur les façades rocheuses faisant face au nord. La plupart des espèces présentes parmi les populations de bryophytes, y compris les lichens, ont été également observées dans ces habitats et plus fréquemment à l'ombre de grosses pierres ou même sous les fissures que celles-ci portent - en particulier au bord des éléments modelés du sol. A une altitude supérieure à 100 mètres, l'aridité augmente et, à des altitudes plus élevées, seuls des Schistidium antarctici (à 500 m dans la vallée Moutonnée) et des Tortella fragilis (près du sommet du pic le plus élevé du sud-ouest, dans la vallée Ablation - 775 m) ont été signalés. Dans ces habitats plus secs, les lichens sont plus fréquents, notamment là où le substrat est stable. Les lichens se répandent et sont localement abondants sur les éboulis, corniches et plateaux les plus stables de la vallée, l'espèce prédominante étant Usnea sphacelata, qui donne une teinte noire aux surfaces rocheuses. Cette espèce se mélange souvent à Pseudephebe minuscula, à plusieurs espèces de lichens encroûtants et, rarement, à Umbilicaria decussata, atteignant le sommet du massif ; à l'exception de cette dernière espèce, toutes poussent aussi régulièrement dans la vallée Moutonnée. Les lichens épiphytes et terricoles, dont les espèces Leproloma cacuminum blanches incrustées prédominent, sont assez répandus là où la surface marginale des bryophytes est plus sèche. D'autres taxons comme Cladonia galindezii, C. pocillum et plusieurs lichens encroûtants sont parfois aussi présents. Divers lichens colonisent le sol sec et les roches de ces lieux, s'étendant parfois sur des coussins de mousses. Il s'agit de Candelariella vitellina, Physcia caesia, Physconia muscigena, et parfois Rhizoplaca melanoplhthalma, Usnea antarctica, Xanthoria elegans, ainsi que de plusieurs espèces non identifiées de taxons encroûtants (notamment les espèces Buellia et Lecidea). Une abondance de Physcia et de Xanthoria dans des endroits isolés semble indiquer l'enrichissement en azote émanant de labbes de McCormick (Stercorarius maccormicki) qui nichent dans la zone (Bentley, 2004). Quelques lichens ornithocoprophiles se développent sur les rochers occasionnels qui servent de perchoirs aux oiseaux. Nombre de bryophytes et lichens se trouvent à la limite australe de leurs répartitions, tandis que plusieurs espèces sont très rares en Antarctique. Les espèces rares de la zone comprennent Bryoerythrophyllum recurvirostrum, Campylium polygamum, Encalypta rhaptocarpa, Tortella alpicola et Tortella fragilis. Plusieurs espèces de Bryum, Encalypta rhaptocarpa, Schistidium occultum et Schistidium chrysoneurum poussent toutes à la limite australe observée pour ces mêmes espèces. Concernant la flore de lichens, la vallée Ablation reste le seul site de l'hémisphère sud où l'on observe la présence de Eiglera flavida et où les Mycobilimbia lobulata et Stereocaulon antarcticum sont également rares. Les espèces de lichens dont les données ont été recueillies à l'extrémité sud sont Cladonia galindezii, Cladonia pocillum, Ochrolechia frigida, Phaeorrhiza nimbosa, Physconia muscigena, et Stereocaulon antarcticum.
Invertébrés, champignons et bactéries
La faune de micro-invertébrés jusqu'ici décrite se fonde sur dix échantillons prélevés dans la vallée Ablation et comprend sept taxons confirmés (Convey et Smith, 1997) : deux collemboles (Cryptopygus badasa, Friesea topo) ; un acarien cryptostigmatide (Magellozetes antarcticus) ; et quatre acariens prostigmatides (Eupodes parvus, Nanorchestes nivalis (= N. gressitti), Rhagidia gerlachei et Stereotydeus villosus). Un certain nombre de spécimens collectés ont été présentés comme Friesea grisea, une espèce très répandue en Antarctique maritime. Néanmoins, les spécimens de Friesia collectés ultérieurement (à partir de 1994) dans l'île Alexandre ont été décrits comme étant une nouvelle espèce différente, F. topo (Greenslade, 1995), elle-même jugée endémique dans l'île Alexandre. Les premières espèces recueillies dans la vallée Ablation ont été réexaminées, toutes celles qui sont restées identifiables étant reclassées comme F. topo. Si le même nombre d'espèces a été décrit sur un autre site de l'île Alexandre, les échantillons en provenance de la vallée Ablation ont affiché une densité totale moyenne de la population de microarthropodes environ sept fois plus importante que celle d'autres sites de la région. La biodiversité dans la vallée Ablation était également supérieure à celle de plusieurs autres sites documentés de l'île Alexandre. Tant la diversité que l'abondance sont considérées comme étant inférieures à celles des sites de la baie Marguerite et plus au nord (Stary et Block, 1998 ; Convey et al., 1996 ; Convey et Smith, 1997 ; Smith, 1996). L'espèce la plus nombreuse enregistrée dans la vallée Ablation était Cryptopygus badasa (96,6 % de tous les arthropodes extraits), particulièrement commune dans les habitats de mousses. L'espèce Friesea topo se rencontrait sur des roches en de faibles densités de populations et était pratiquement absente dans l'habitat des mousses, indiquant que ces espèces devaient avoir des préférences distinctes en matière d'habitat. La vallée Ablation est le seul site de l'île Alexandre où l'on a décrit l'acarien prédateur R. gerlachei. Très peu de recherches ont été menées sur les champignons de la zone ; toutefois, une étude a signalé un champignon attrape-nématodes qui reste à identifier et qui se développe dans un étang de la vallée Ablation (Maslen, 1982). Si un autre échantillonnage est nécessaire pour décrire in extenso la microfaune terrestre, il n'en demeure pas moins que les données disponibles soutiennent l'importance biologique de la zone.
Oiseaux nicheurs
La faune avicole du massif de la vallée Ablation - mont Ganymède n'a pas encore fait l'objet d'une étude détaillée. Quelques couples de labbes de McCormick (Stercorarius maccormicki) ont été signalés près de certains sites à végétation humides (Smith, 1988). Des pétrels des neiges « se reproduiraient » dans les environs de la pointe Ablation (Croxall et al., 1995, citant Fuchs et Adie, 1949). Bentley (2004) a rapporté l'existence d'une prédation aérienne directe commise par des labbes de McCormick sur les pétrels des neiges dans la zone. Aucune autre espèce avicole n'a été enregistrée dans le massif de la vallée Ablation - mont Ganymède.
Activités humaines et leurs impacts
L'activité humaine dans le massif de la vallée Ablation - mont Ganymède est menée à des fins exclusivement scientifiques. La première visite de la vallée Ablation a été réalisée en 1936 par des membres de la British Graham Land Expedition, qui ont prélevé environ 100 spécimens de fossiles non loin de la pointe Ablation (Howlett, 1988). D'autres expéditions ont été effectuées une dizaine d'années plus tard, lorsque des descriptions géologiques de base ainsi que d'autres collectes de fossiles ont été faites. Des recherches paléontologiques plus intensives seront menées par des géologues britanniques entre les années 1960 et 1980, donnant lieu à des études géomorphologiques détaillées (Clapperton et Sugden, 1983). Les études limnologiques ont commencé dans les années 1970 et plusieurs expéditions se sont intéressées à la biologie terrestre dans les années 1980 et 1990. Les activités scientifiques entreprises depuis le début de ce siècle se sont concentrées sur la recherche paléo-climatologique. Toutes les expéditions connues dans la zone ont été effectuées par des scientifiques britanniques. Les impacts de ces activités n'ont pas été décrits de manière détaillée, mais ils semblent mineurs et se limitent à des empreintes de pas, aux traces d'aéronefs sur la piste d'atterrissage de la vallée Moutonnée (voir section 6 [ii] ), à l'extraction de petites quantités d'échantillons géologiques et biologiques, aux bornes, aux objets abandonnés tels que le matériel d'approvisionnement et scientifique, ainsi qu'aux restes de déchets humains.
Un dépôt abandonné, comprenant deux cuves de pétrole (l'une pleine et l'autre vide), trois bidons de 5 litres d'huile pour motoneige, une caisse de nourriture et dix sondes de glacier, se trouvaient sur le banc de moraine adjacent à la plateforme George VI, à quelque 500 mètres au nord du lac Moutonnée (70°51'19" S ; 68°19'05" O). Ce dépôt a été partiellement enlevé en novembre 2012 et les deux cuves pleines de pétrole ont été enlevées en novembre 2013. Plusieurs expéditions effectuées dans les années 1970 et 1980 déposaient des cuves de pétrole vides comme jalons d'itinéraire, à travers la glace de pression, du détroit George VI jusqu'à la vallée Ablation, ainsi qu'une énorme roche de rivage peinte en jaune laissée au sud-est du lac Ablation (McAra, 1984 ; Hodgson, 2001). Dans les environs, se trouve une grosse croix faite de roches et de cairns peints en rouge, avec un tableau de marquage en bois placé en son centre. Des éléments indiquant les sites de campement sur la rive du lac Ablation étaient encore visibles en 2012. Un site se trouve sur la rive sud-ouest, près d'une zone végétale riche, et un autre se trouve à quelque quatre kilomètres à l'est sur la rive sud-est. Sur les deux sites, des cercles de pierres marquent les emplacements de vieilles tentes, alors que des structures circulaires ont été construites avec des murs de pierre bas (0,8 m). Sur l'ancien site, nombre de pièces en bois (comme de vieilles bornes), une vieille caisse de nourriture, du fil et des déchets humains ont été observés (Harris, 2001 ; Hodgson, 2001). On a trouvé plusieurs roches peintes en rouge autour des rives sud et ouest du lac Ablation en février 2001, ainsi que des fragments de peinture dans des sédiments. En 2000-2001, certains des objets abandonnés dans la vallée Ablation ont été enlevés. Il s'agit de trois cuves de pétrole trouvées sur la glace du lac, d'une vieille caisse de nourriture, de bois et de ficelles trouvés sur la rive sud-ouest, ainsi que de nombreux fragments issus de cloches acryliques en plexiglas brisées récupérés sur la rive sud-ouest (neuf ont été enlevés en janvier 1993 - Wynn-Williams, 1993 ; Rossaak, 1997 - tous étaient détruits par le vent) (Harris, 2001 ; Hodgson, 2001). En novembre 2012, du métal et des déchets ont été enlevés près d'un vieux campement ayant un mur de pierres bas (situé à 70°49'58" S ; 68°22'16" O). Les roches peintes demeurent. Des motoneiges ont été utilisées sur le lac gelé et le glacier et des motoneiges adaptées - avec des roues avant, ont été utilisées sur le terrain en gravier à une proximité réduite de la rive sud-ouest du lac Ablation en 1983 et 1984 (McAra, 1984). Quelques traces des passages érodés sur des pentes raides en éboulis, probablement du fait du travail de terrain, ont été enregistrées dans la vallée Moutonnée (Howlett, 1988). Des cairns ont été construits sur plusieurs sommets montagneux afin de marquer des sites de prélèvements à travers la zone.
6 (ii) Accès à la zone
L'accès à la zone se fait par avion, véhicule ou à pied.
Il n'y a pas de restrictions spéciales en ce qui concerne les points d'accès à et de départ de la zone, que ce soit par voie terrestre ou par voie aérienne. L'accès par voie terrestre à partir de la plateforme George VI peut se révéler difficile en raison de la glace de pression, mais il est considéré comme l'accès le plus fiable et le plus sûr pour les visiteurs qui arrivent dans les environs de la zone par aéronef à voilure fixe, notamment parce que certains itinéraires reliant les glaciers à la zone sont accidentés, crevassés et ardus.
Il est déconseillé d'atterrir avec des aéronefs à voilure fixe dans la zone. Si des impératifs de recherche scientifique ou de gestion imposent un atterrissage, il doit se limiter aux lacs recouverts de glace ou à un seul site terrestre situé directement à l'ouest du lac Moutonnée et à condition que l'atterrissage soit faisable. La déformation par pression de la surface glaciaire des lacs, de l'eau de fonte et de la couverture glaciaire de mince épaisseur peut rendre impraticable tout atterrissage sur le lac à la fin de l'été. Des atterrissages sur le lac Ablation et sur le site terrestre situé à l'ouest du lac Moutonnée ont été effectués en novembre 2000. Le site d'atterrissage terrestre (carte 3) est orienté est-ouest avec près de 350 mètres de piste revêtue de graviers et à pente douce relevée à environ 2 mètres au-dessus de la vallée environnante. Quelques pierres peintes en rouge marquent l'extrémité (supérieure) ouest, sous la forme d'une flèche. Des empreintes de pneus sont visibles sur les graviers. En raison du mauvais état de la piste et du risque de dommages pour l'aéronef, il n'est pas recommandé d'utiliser le site terrestre situé à l'ouest du lac Moutonnée.
Si l'accès par hélicoptère se révèle faisable, aucun site spécifique d'atterrissage n'a été désigné, mais atterrir à 200 mètres des rives du lac, ou à 100 mètres de tout sol humide ou à couvert végétal, ou même sur des lits de cours d'eau, est interdit.
L'accès est également possible par voie aérienne sur la partie supérieure du glacier Jupiter (550 m), immédiatement à l'ouest de la vallée Ablation et hors de la zone, lieux à partir desquels l'accès à la zone peut se faire par voie terrestre à pied.
Il est interdit aux pilotes, membres d'équipage et à toute autre personne arrivant par voie aérienne de se déplacer à pied au-delà des alentours immédiats de tout site d'atterrissage de la zone, sauf si un permis les y autorise spécifiquement.
6 (iii) Structures à l'intérieur et à proximité de la zone
La zone n'abrite aucune structure connue. Quelques cairns ont été posés pour servir de bornes lors des levés topographiques dans toute la zone (Perkins, 1995, Harris, 2001) et les sites de campement disposent de quelques murs de faible hauteur. Neuf balises en plastique de couleur rouge vif (de 30 cm de haut et retenues au sol à l'aide de pierres) ont été installées pour délimiter la piste d'atterrissage dans la vallée Moutonnée, avant d'être retirées en novembre 2012. La structure la plus proche de la zone semble être un refuge abandonné dans le Spartan Cwm à environ 20 km au sud de la zone. Un seul campement estival réservé à la recherche scientifique existe à Fossil Bluff (Royaume-Uni), à près de 60 km au sud de la côte est de l'île Alexandre. Les stations de recherche scientifique permanentes les plus proches se trouvent à la baie Marguerite. Il s'agit des stations General San Martin (Argentine) et Rothera Research Station (Royaume-Uni) situées à environ 350 km plus au nord (carte 2).
6 (iv) Emplacement d'autres zones protégées à proximité
Il n'existe pas d'autres zones protégées dans les environs immédiats de la zone. La zone protégée la plus proche de la vallée Ablation et mont Ganymède est la ZSPA n° 170 : Nunataks Marion, île Charcot, péninsule antarctique à environ 270 km à l'est de l'île Alexandre (carte 2).
6 (v) Zones spéciales à l'intérieur de la zone
Il n'y a aucune zone spéciale à l'intérieur de la zone.
7. Critères de délivrance des permis
7 (i) Conditions générales pour l'obtention d'un permis
L'accès à la zone est interdit sauf avec un permis délivré par une autorité nationale compétente. Les critères de délivrance d'un permis pour entrer dans la zone sont les suivants :
- un permis est délivré pour des raisons scientifiques indispensables qu'il est impossible de satisfaire ailleurs ou pour des raisons de gestion essentielles à la zone ;
- les activités autorisées sont conformes au présent plan de gestion ;
- toutes les activités de gestion soutiennent la réalisation des buts et objectifs du présent plan de gestion ;
- les activités autorisées ne mettront pas en péril l'écosystème naturel de la zone ;
- les activités autorisées veilleront, au moyen d'un processus d'évaluation d'impact sur l'environnement, à la protection permanente des valeurs environnementales et scientifiques de la zone ;
- le permis est délivré pour une période limitée ;
- le permis, ou une copie certifiée, sera emporté à l'intérieur de la zone.
7 (ii) Accès à la zone et déplacements à l'intérieur de celle-ci
Les mouvements de véhicules à l'intérieur de la zone ne peuvent s'effectuer que sur des surfaces de neige ou de glace.
Les mouvements terrestres à l'intérieur de la zone doivent se faire à pied.
Tout mouvement doit se faire avec la plus grande précaution afin de réduire au minimum toute perturbation du sol, des couverts végétaux et des éléments géomorphologiques sensibles tels que les dunes ; marcher sur la neige ou sur des terrains rocailleux, si possible. Si possible, les visiteurs doivent éviter de marcher sur les cours d'eau ou sur les lits de lacs secs, ou même sur le sol humide, afin de ne pas perturber l'hydrologie et/ou endommager les communautés végétales sensibles. Il convient de faire attention même lorsqu'il n'y a pas d'humidité visible, puisque les plantes discrètes peuvent toujours coloniser le sol.
Les déplacements à pied doivent être limités au minimum requis pour effectuer les activités autorisées et tous les efforts raisonnables doivent être consentis pour réduire les effets du piétinement.
Les opérations de survol des zones doivent être réalisées conformément aux « Directives pour l'exploitation d'aéronefs à proximité de concentrations d'oiseaux dans l'Antarctique », inscrites dans la résolution 2 (2004).
Le survol de colonies d'oiseaux dans la zone par des systèmes d'aéronefs pilotés à distance (RPAS) n'est pas autorisé, sauf à des fins scientifiques ou opérationnelles, et en vertu d'un permis émis par une autorité nationale compétente.
7 (iii) Activités pouvant être conduites à l'intérieur de la zone
Les activités pouvant être menées dans la zone sont :
- les activités de gestion essentielles, y compris de suivi ;
- des travaux de recherche scientifique indispensables qui ne peuvent être entrepris ailleurs et ne risquent pas de mettre en péril l'écosystème de la zone ; et
- l'échantillonnage qui doit être réduit au minimum pour répondre aux programmes de recherches approuvés.
En temps normal, effectuer des plongées dans les lacs de la zone est interdite, sauf si elle se justifie par des considérations scientifiques impérieuses. En cas de plongée, une attention particulière doit être de mise afin de ne pas perturber la colonne d'eau, les sédiments et les communautés biologiques sensibles. Il conviendra de tenir compte, lors de la délivrance des permis de plongée, de la sensibilité de la colonne d'eau, des sédiments et des communautés biologiques aux perturbations imputables à telles activités.
7 (iv) Installation, modification ou enlèvement de structures
Les structures ou installations permanentes sont interdites.
Aucune structure ne doit être érigée dans la zone et aucun matériel scientifique ne doit y être installé, sauf pour des raisons scientifiques ou de gestion indispensables et pour une période préétablie définies dans un permis.
Toutes les bornes, les structures et tout l'équipement scientifique installés dans la zone doivent clairement identifier le pays, le nom du principal chercheur ou de la principale agence, l'année d'installation et la date d'enlèvement prévue.
Ces éléments ne doivent porter aucun organisme, propagule (semences, œufs, spores, etc.) ou sol non stérile (voir section 7 [vi] ). Les éléments doivent être composés de matériaux pouvant résister aux conditions environnementales et de ce fait réduire au minimum le risque de contamination de la zone.
L'enlèvement de structures et matériels particuliers, une fois le permis les autorisant expiré, relèvera de la responsabilité de l'autorité qui a délivré le permis initial ; le permis doit être assorti de cette condition.
7 (v) Emplacement des camps
Lorsque certaines opérations prévues dans le permis l'exigent, des camps temporaires peuvent être installés dans la zone. Un site aménagé à cet effet est situé à l'extrémité nord-ouest (supérieure) de la piste d'atterrissage dans la vallée Moutonnée (70°51'48" S, 68°21'39" O) (carte 3). Bien que ce site ne soit pas balisé, les tentes doivent être installées le plus près possible de la balise située à l'extrémité nord-ouest de la piste d'atterrissage. Il est toujours préférable d'utiliser ce site lorsque des activités doivent être menées dans les alentours. A ce jour, aucun autre emplacement n'a été désigné pour l'installation de camps, mais tout campement est de toute manière interdit aux endroits abritant de la végétation. En outre, tout campement doit être situé le plus loin possible (au moins 200 m de préférence) des berges des lacs ainsi que des lits asséchés de lacs et de cours d'eau (qui peuvent abriter un biote invisible à l'œil nu). Tout camp doit être de préférence installé, lorsque la situation le permet, sur des surfaces enneigées ou couvertes de glace. Tout emplacement préalablement utilisé à cette fin doit être réutilisé dans la mesure du possible sauf si, compte tenu des critères susmentionnés, ces emplacements se révèlent inappropriés.
7 (vi) Restrictions relatives aux matériaux et organismes pouvant être introduits dans la zone
L'introduction délibérée dans la zone d'animaux, de végétaux ou de micro-organismes vivants est interdite. Pour assurer la conservation des valeurs écologiques de la zone, des précautions particulières doivent être prises contre l'introduction accidentelle de microbes, invertébrés ou plantes provenant d'autres sites de l'Antarctique y compris les stations ou des régions extérieures à l'Antarctique. Tout le matériel d'échantillonnage et les balises introduits dans la zone doivent être nettoyés et stérilisés. Dans la mesure du possible, les chaussures et autres équipements utilisés ou introduits dans la zone (y compris les sacs et les sacs à dos) doivent être minutieusement nettoyés avant d'entrer dans la zone. Des conseils supplémentaires sont donnés dans le Manuel sur les espèces non indigènes du CPE (édition 2017) et l'Environmental code of conduct for terrestrial scientific field research in Antarctica [Code de conduite environnementale pour la recherche scientifique sur le terrain en Antarctique] (SCAR 2009). Compte tenu de la présence éventuelle de colonies d'oiseaux nicheurs dans la zone, aucun produit provenant ou dérivé d'espèces avicoles - notamment les déchets, les produits contenant des œufs en poudre non pasteurisés - ne doit être introduit dans la zone.
Aucun herbicide ni pesticide ne doit être introduit dans la zone. Tout autre produit chimique, y compris les radionucléides ou les isotopes stables, qui peuvent être introduits pour des raisons scientifiques ou des raisons de gestion visées dans le permis, seront enlevés de la zone au plus tard à la fin de l'activité pour laquelle le permis a été délivré. L'émission, directement dans l'environnement, de radionucléides ou d'isotopes stables par une méthode les rendant irrécupérables doit être évitée. Ni le carburant, ni tout autre matériau chimique, ne peuvent être entreposés dans la zone, à moins que le permis ne l'autorise spécifiquement. Ils doivent être stockés et manipulés de façon à limiter le risque d'introduction accidentelle dans l'environnement. Les matériaux introduits dans la zone le sont pour une période donnée uniquement et doivent être enlevés lors de, ou avant la conclusion de ladite période. En cas de déversement susceptible de mettre en péril les valeurs de la zone, leur retrait est encouragé à condition que l'impact de celui-ci ne soit pas susceptible d'être supérieur à celui consistant à laisser les substances in situ. L'autorité compétente doit être informée de tout élément introduit et non retiré qui ne figurait pas dans le permis agréé.
7 (vii) Prélèvement de végétaux, capture d'animaux ou perturbations nuisibles de la faune et la flore
Toute capture d'animaux ou toute perturbation nuisible à la faune et la flore indigène est interdite sauf avec un permis distinct délivré spécifiquement à cette fin en vertu de l'annexe II du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement. Dans le cas de prélèvements ou de perturbations nuisibles aux animaux, le SCAR code of conduct for the use of animals for scientific purposes in Antarctica [Code de conduite du SCAR pour l'utilisation d'animaux à des fins scientifiques dans l'Antarctique] (2011) devra être utilisé comme norme minimale. Les activités d'échantillonnage du sol ou de la végétation doivent être limitées à leur strict minimum requis à des fins scientifiques ou à des fins de gestion et exécutées avec des techniques qui minimisent les perturbations susceptibles d'être causées au sol, aux structures de glace et au biote.
7 (viii) Collecte ou retrait de matériaux non introduits dans la zone par le titulaire du permis
Le ramassage ou l'enlèvement de matériaux dans la zone ne peut se faire que sur délivrance d'un permis et doivent se limiter au minimum requis pour les activités menées à des fins scientifiques ou de gestion. Les matériaux d'origine humaine susceptibles de mettre en péril les valeurs de la zone, qui n'ont pas été introduits dans celle-ci par le détenteur du permis ou qui n'ont pas été autrement autorisés, peuvent être enlevés de la zone à moins que l'impact environnemental de l'enlèvement ne soit plus grand que si les matériaux sont laissés in situ. Si tel est le cas, l'autorité compétente doit en être informée et son autorisation obtenue.
7 (ix) Elimination des déchets
Tous les déchets, à l'exception des déchets liquides domestiques et humains, seront retirés de la zone. Les déchets liquides domestiques et humains peuvent être éliminés de la zone en les évacuant dans les failles de glace longeant le bord de la plateforme glaciaire George VI ou du glacier Jupiter, ou en les enterrant dans la moraine située le long de la marge glaciaire à ces mêmes endroits, le plus près possible de la glace.
Toute élimination des déchets liquides domestiques et humains selon cette méthode devra s'effectuer à une distance minimum de 200 mètres et impérativement hors des zones de captation des principaux lacs des vallées Ablation, Moutonnée et Flatiron. Si ces conditions ne peuvent être satisfaites, le déchet doit être retiré autrement de la zone. Les déchets humains solides doivent être enlevés de la zone.
7 (x) Mesures qui peuvent être nécessaires pour continuer de répondre aux objectifs du plan de gestion
Des permis peuvent être délivrés pour entrer dans la zone afin d'y mener des travaux de recherche scientifique, de surveillance et d'inspection de site, qui impliquent le prélèvement d'un petit nombre d'échantillons à des fins d'analyse ou pour appliquer des mesures de protection.
Tous les sites de suivi de longue durée doivent être signalés par des bornes ou des panneaux dûment entretenus.
Les activités scientifiques devront être menées conformément au Environmental code of conduct for terrestrial scientific field research in Antarctica [Code de conduite en matière environnementale pour la recherche scientifique terrestre sur le terrain dans l'Antarctique] (SCAR, 2009).
7 (xi) Rapports de visite
Le principal détenteur du permis soumettra, pour chaque visite dans la zone, un rapport à l'autorité nationale compétente, dès que possible, et au plus tard six mois après la fin de ladite visite. Ces rapports doivent contenir, le cas échéant, les catégories d'informations mentionnées dans le formulaire de rapport de visite repris dans le Guide révisé pour l'élaboration des plans de gestion pour les zones spécialement protégées de l'Antarctique (annexe 2). L'autorité nationale doit, chaque fois qu'elle le peut, également transmettre une copie du rapport de visite à la Partie étant à l'initiative du plan de gestion, afin de l'aider à la gestion de la zone et dans la révision du plan de gestion. Les Parties doivent, dans la mesure du possible, déposer les originaux ou des copies de ces rapports dans une archive à laquelle le public pourra avoir accès afin de maintenir ainsi une archive d'usage. Cette archive pourra être utilisée à la fois lors de tout réexamen du plan de gestion et dans le cadre de l'organisation de l'utilisation scientifique de la zone.
8. Bibliographie
Bell, C. M. (1975). Structural geology of parts of Alexander Island. British Antarctic Survey Bulletin 41 et 42 : 43-58.
Bentley, M. J. (2004). Aerial predation by a south polar skua Catharacta maccormicki on a snow petrel Pagodroma nivea in Antarctica. Marine Ornithology 32 : 115-116.
Bentley, M. J., Hodgson, D. A., Sugden, D. E., Roberts, S. J., Smith, J. A., Leng, M. J., Bryant, C. (2005). Early Holocene retreat of George VI Ice Shelf, Antarctic Peninsula. [Retraite holocène précoce de la plate-forme de glace George VI, péninsule Antarctique.] Géologie 33 : 173-176.
Bentley, M. J., Hodgson, D. A., Smith, J. A., Cofaigh, C. O., Domack, E. W., Larter, R. D., Roberts, S. J., Brachfeld, S., Leventer, A., Hjort, C., Hillenbrand, C. D., and Evans, J. (2009). Mechanisms of Holocene palaeoenvironmental change in the Antarctic Peninsula region. [Mécanismes de changement paléoenvironnemental Holocène dans la région de la péninsule Antarctique.] The Holocene 19 : 51-69.
Butterworth, P. J. (1985). Sedimentology of Ablation Valley, Alexander Island : report on Antarctic field work. British Antarctic Survey Bulletin 66 : 73-82.
Butterworth, P. J., Crame, J. A., Howlett, P. J., et Macdonald, D. I. M. (1988). Lithostratigraphy of Upper Jurassic - Lower Cretaceous strata of eastern Alexander Island, Antarctica. [Lithostratigraphie du Jurassique supérieur - Crétacé inférieur de l'est de l'île Alexandre, de l'Antarctique.] Cretaceous Research 9 : 249-64.
Clapperton, C. M. , and Sugden, D. E. (1982). Late Quaternary glacial history of George VI Sound area, West Antarctica. Quaternary Research 18 : 243-67.
Clapperton, C. M. , and Sugden, D. E. (1983). Geomorphology of the Ablation Point massif, Alexander Island, Antarctica. Boreas 12 : 125-35.
Comité pour la protection de l'environnement (CPE) (2017). Manuel des espèces non indigènes - 2e édition. Manuel préparé par le Groupe de contact intersessions du CPE et adopté par la Réunion Consultative du Traité sur l'Antarctique par la résolution 4 (2016). Buenos Aires, Secrétariat du Traité sur l'Antarctique.
Convey, P., Greenslade, P., Richard, K. J., et Block W. (1996). The terrestrial arthropod fauna of the Byers Peninsula, Livingston Island, South Shetland Islands - Collembola. Polar Biology 16 : 257-59.
Convey, P., and Smith, R. I. L. (1997). The terrestrial arthropod fauna and its habitats in northern Marguerite Bay and Alexander Island, maritime Antarctic. Antarctic Science 9 : 12-26.
Crame, J. A. (1981). The occurrence of Anopaea (Bivalvia : Inoceramidae) in the Antarctic Peninsula. Journal of Molluscan Studies 47 : 206-219.
Crame, J. A. (1985). New Late Jurassic Oxytomid bivalves from the Antarctic Peninsula region. British Antarctic Survey Bulletin 69 : 35-55.
Crame, J. A., et Howlett, P. J. (1988). Late Jurassic and Early Cretaceous biostratigraphy of the Fossil Bluff Formation, Alexander Island. British Antarctic Survey Bulletin 78 : 1-35.
Croxall, J. P., Steele, O. K., McInnes, S. J., et Prince, P. A. (1995). Breeding distribution of the snow petrel Pagodroma nivea. Marine Ornithology 23 : 69-99.
Elliott, M. R. (1974). Stratigraphy and sedimentary petrology of the Ablation Point area, Alexander Island. British Antarctic Survey Bulletin 39 : 87-113.
Greenslade, P. (1995). Collembola from the Scotia Arc and Antarctic Peninsula including descriptions of two new species and notes on biogeography. Polskie Pismo Entomologiczne 64 : 305-19.
Barton C. M. 2001. Révision des plans de gestion pour les zones protégées de l'Antarctique proposés à l'origine par les États-Unis d'Amérique et le Royaume-Uni : Rapport de visite sur le terrain. Rapport interne
pour la National Science Foundation, US, et le Foreign and Commonwealth Office, RU. Environmental Research and Assessment, Cambridge.
Heywood, R. B. (1977). A limnological survey of the Ablation Point area, Alexander Island, Antarctica. Philosophical Transactions of the Royal Society B 279 : 39-54.
Heywood, R. B., et Light, J. J. (1975). First direct evidence of life under Antarctic shelf ice. Nature 254: 591-92.
Hodgson, D. 2001. Millennial-scale history of the George VI Sound ice shelf and palaeoenvironmental history of Alexander Island. Rapport scientifique de la BAS - Sledge Charlie 2000-2001. Ref. R/2000/NT5.
Howlett, P. J. (1986). Olcostephanus (Ammonitina) from the Fossil Bluff Formation, Alexander Island, and its stratigraphical significance. British Antarctic Survey Bulletin 70 : 71-77.
Howlett, P. J. (1988). Latest Jurassic and Early Cretaceous cephalopod faunas of eastern Alexander. Island, Antarctica. Thèse de doctorat non publiée, University College, Londres.
Light, J. J., et Heywood, R. B. (1975). Is the vegetation of continental Antarctica predominantly aquatic ? Nature 256 : 199-200.
Lipps, J. H., Krebs, W. N., and Temnikow, N. K. (1977). Microbiota under Antarctic ice shelves. Nature 265 : 232-33.
Maslen, N. R. (1982). An unidentified nematode-trapping fungus from a pond on Alexander Island. British Antarctic Survey Bulletin 51 : 285-87.
Morgan, F., Barker, G., Briggs, C. Price, R., et Keys, H. (2007). Environmental Domains of Antarctica Version 2.0 Final Report. Landcare Research Contract Report LC0708/055.
Roberts, S. J., Hodgson, D. A., Bentley, M. J., Smith, J. A., Millar, I. L., Olive, V., et Sugden, D. E. (2008). The Holocene history of George VI Ice Shelf, Antarctic Peninsula from clast-provenance analysis of epishelf lake sediments. Palaeogeography, Palaeoclimatology, Palaeoecology 259 : 258-283.
Roberts, S. J., Hodgson, D. A., Bentley, M. J., Sanderson, D. C. W., Milne, G., Smith, J. A., Verleyen, E., and Balbo, A. (2009). Holocene relative sea-level change and deglaciation on Alexander Island, Antarctic Peninsula, from elevated lake deltas. Geomorphology 112 : 122-134.
P. D., Rowley et Smellie, J. L. (1990). Southeastern Alexander Island. Dans : LeMasurier, W. E., et Thomson, J. W., eds. Volcanoes of the Antarctic plate and southern oceans. Antarctic Research Series 48. Washington D.C., American Geophysical Union 277-279.
SCAR (Comité scientifique pour la recherche en Antarctique) (2009) Environmental code of conduct for terrestrial scientific field research in Antarctica [Code de conduite environnementale pour la recherche scientifique sur le terrain en Antarctique]. XXXIIe RCTA IP4.
SCAR (Comité scientifique pour la recherche en Antarctique) (2011) SCAR code of conduct for the use of animals for scientific purposes in Antarctica [Code de conduite du SCAR pour l'utilisation d'animaux à des fins scientifiques]. XXXIIe RCTA IP53.
Smith, J. A., Bentley, M. J., Hodgson, D. A., Roberts, S. J., Leng, M. J., Lloyd, J. M. , Barrett, M. S., Bryant, C., et Sugden, D. E. (2007 a). Oceanic and atmospheric forcing of early Holocene ice shelf retreat, George VI Ice Shelf, Antarctica Peninsula. Quaternary Science Reviews 26 : 500-516.
Smith, J. A., Bentley, M. J., Hodgson, D. A., and Cook, A. J. (2007b) George VI Ice Shelf : past history, present behaviour and potential mechanisms for future collapse. Antarctic Science 19 : 131-142.
Smith, R. I. L. (1988). Bryophyte oases in ablation valleys on Alexander Island, Antarctica. The Bryologist 91 : 45-50.
Smith, R. I. L. (1996). Terrestrial and freshwater biotic components of the western Antarctic Peninsula.
Dans : In Ross, R.M., Hofmann, E.E. and Quetin, L.B. (eds) Foundations for ecological research west of the Antarctic Peninsula. Antarctic Research Series 70 : American Geophysical Union, Washington D.C. : 15-59.
Smith, R. I. L. (1997). Oases as centres of high plant diversity and dispersal in Antarctica. Dans : Lyons, W.B., Howard-Williams, C. & Hawes, I. (Eds) Ecosystem processes in Antarctic ice-free landscapes. A.A. Balkema, Rotterdam : 119-28.
Smith, R. I. L., and Convey, P. (2002). Enhanced sexual reproduction in bryophytes at high latitudes in the maritime Antarctic. Journal of Morphology 24 : 107-117.
Starý, J., and Block, W. (1998). Distribution and biogeography of oribatid mites (Acari : Oribatida) in Antarctica, the sub-Antarctic and nearby land areas. Journal of Natural History 32 : 861-94.
Sugden, D. E., et Clapperton, C. N. (1980). West Antarctic ice sheet fluctuations in the Antarctic Peninsula area. Nature 286 : 378-81.
Sugden, D. E., et Clapperton, C. M. (1981). An ice-shelf moraine, George VI Sound, Antarctica. Annals of Glaciology 2 : 135-41.
Taylor, B. J., Thomson, M. R. A., and Willey, L. E. (1979). The geology of the Ablation Point - Keystone Cliffs area, Alexander Island. British Antarctic Survey Scientific Report 82.
Terauds, A., and Lee, J. R. (2016). Antarctic biogeography revisited : updating the Antarctic Conservation Biogeographic Regions. Diversity and Distribution 22 : 836-840.
Terauds, A., Chown, S. L., Morgan, F., Peat, H. J., Watt, D., Keys, H., Convey, P., et Bergstrom, D. M. (2012). Conservation biogeography of the Antarctic. Diversité et Distribution 18 : 726-41.
Thomson, M. R. A. (1972). Ammonite faunas of south-eastern Alexander Island and their stratigraphical significance. Dans : Adie, R.J. (ed) Antarctic Geology and Geophysics, Universitetsforlaget, Oslo.
Thomson, M. R. A. (1979). Upper Jurassic and Lower Cretaceous Ammonite faunas of the Ablation Point area, Alexander Island. British Antarctic Survey Scientific Report 97.
Thomson, M. R. A., et Willey, L. E. (1972). Upper Jurassic and Lower Cretaceous Inoceramus (Bivalvia) from south-east Alexander Island. British Antarctic Survey Bulletin 29 : 1-19.
Willey, L. E. (1973). Belemnites from south-eastern Alexander Island : II The occurrence of the family Belemnopseidae in the Upper Jurassic and Lower Cretaceous. British Antarctic Survey Bulletin 36 : 33-59.
Willey, L. E. (1975). Upper Jurassic and Lower Cretaceous Pinnidae (Bivalvia) from southern Alexander Island. British Antarctic Survey Bulletin 41 & 42 : 121-31.
Carte 1. - Coordonnées de la vallée Ablation et mont Ganymède, dans la péninsule antarctique. Spécifications de la carte : WGS84 Stéréographique polaire antarctique. Méridien central : - 55° : Parallèle de référence : - 71°.
Carte 2. - Coordonnées de la ZSPA n° 147, vallée Ablation et mont Ganymède, île Alexandre Spécifications de la carte : WGS 1984 Stéréographique polaire antarctique. Méridien central : - 71° ; Parallèle de référence : - 71°.
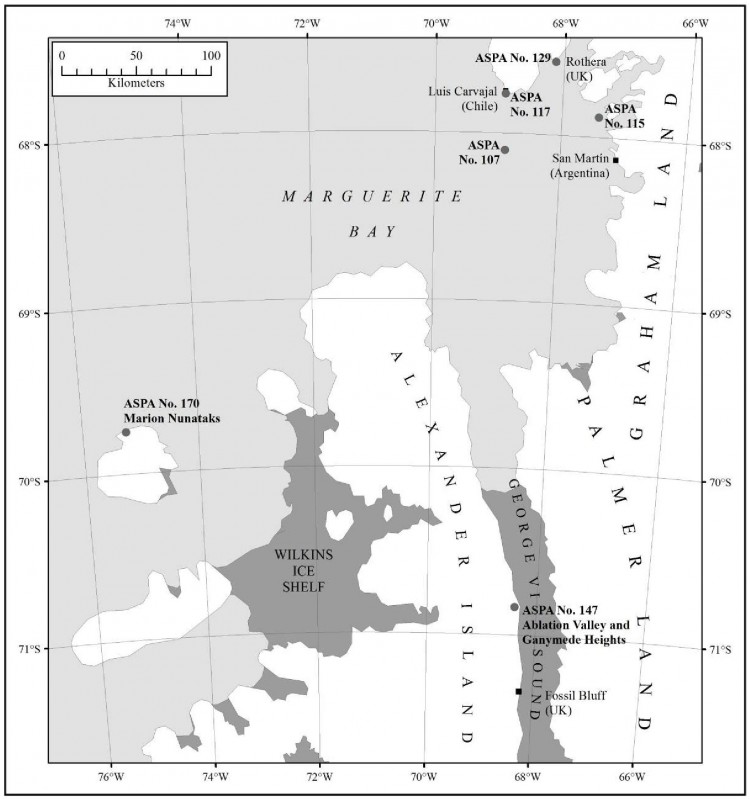
Carte 3. - Carte topographique de la ZSPA n° 147, vallée Ablation et mont Ganymède, île Alexandre Spécifications de la carte : WGS 1984 Stéréographique polaire antarctique. Méridien central : - 68,4° ; Parallèle de référence : - 71,0°.
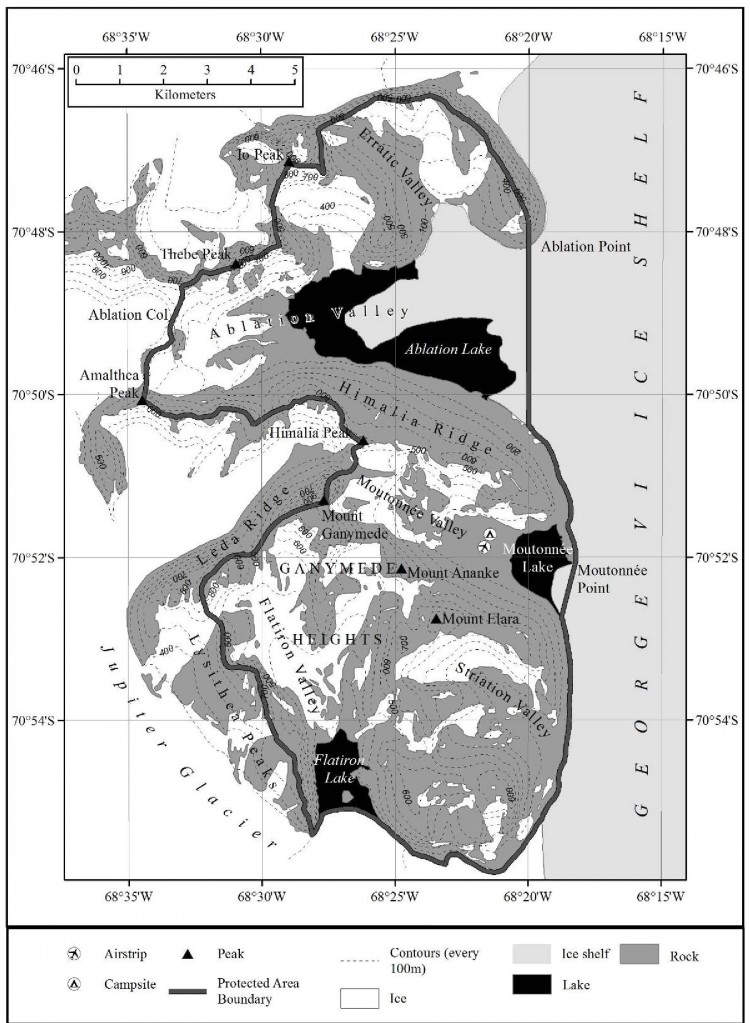
ANNEXE 1
Coordonnées des limites de la ZSPA n° 147, vallée Ablation et mont Ganymède, île Alexandre. En grande partie, la limite suit les caractéristiques naturelles et une description détaillée se trouve dans la section 6 (i). Dans le tableau ci-dessous, les coordonnées des limites sont numérotées, le numéro 1 correspondant aux coordonnées les plus septentrionales et les suivantes étant numérotées de manière séquentielle dans le sens des aiguilles d'une montre autour des limites de la zone.
|
Nombre |
Latitude |
Longitude |
|
1 |
70°46'26”S |
68°24'01”O |
|
2 |
70°46'28”S |
68°25'48”O |
|
3 |
70°46'55”S |
68°28'27”O |
|
4 |
70°47'13”S |
68°28'15”O |
|
5 |
70°47'12”S |
68°29'33”O |
|
6 |
70°48'02”S |
68°29'58”O |
|
7 |
70°48'23”S |
68°32'55”O |
|
8 |
70°49'44”S |
68°34'38”O |
|
9 |
70°50'06”S |
68°31'13”O |
|
10 |
70°49'56”S |
68°28'52”O |
|
11 |
70°50'19”S |
68°26'51”O |
|
12 |
70°51'17”S |
68°28'19”O |
|
13 |
70°52'09”S |
68°31'59”O |
|
14 |
70°53'02”S |
68°31'06”O |
|
15 |
70°53'03”S |
68°29'59”O |
|
16 |
70°55'03”S |
68°27'58”O |
|
17 |
70°54'53”S |
68°27'40”O |
|
18 |
70°55'36”S |
68°23'26”O |
|
19 |
70°55'41”S |
68°21'30”O |
|
20 |
70°54'43”S |
68°19'11”O |
|
21 |
70°52'44”S |
68°19'03”O |
|
22 |
70°52'04”S |
68°18'25”O |
|
23 |
70°51'17”S |
68°18'41”O |
|
24 |
70°50'18”S |
68°20'27”O |
|
25 |
70°48'08”S |
68°20'44”O |
|
26 |
70°47'38”S |
68°21'23”O |
|
27 |
70°46'55”S |
68°22'16”O |
Annexe
MESURE 5 (2018)
ZONE SPÉCIALEMENT PROTÉGÉE DE L'ANTARCTIQUE N° 170 (NUNATAKS MARION, ILE CHARCOT, PÉNINSULE ANTARCTIQUE) : PLAN DE GESTION RÉVISÉ (ENSEMBLE UNE ANNEXE)
Les Représentants,
Rappelant les articles 3, 5 et 6 de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement qui disposent de la désignation des zones spécialement protégées de l'Antarctique (« ZSPA ») et de l'adoption des plans de gestion pour ces zones ;
Rappelant
- la mesure 4 (2008) qui désignait les nunataks Marion, l'île Charcot, la péninsule antarctique comme ZSPA n° 170 et adoptait un plan de gestion pour cette zone ;
- la mesure 16 (2013), qui a adopté un plan de gestion révisé pour la ZSPA n° 170 ;
Notant que le Comité pour la protection de l'environnement a approuvé un plan de gestion révisé pour la ZSPA n° 170 ;
Souhaitant remplacer le plan de gestion actuel pour la ZSPA n° 170 par le plan de gestion révisé ;
Recommandent à leurs gouvernements d'approuver la mesure suivante conformément au paragraphe 1 de l'article 6 de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement :
Que :
1. Le plan de gestion révisé pour la zone spécialement protégée de l'Antarctique n° 170 (Nunataks Marion, île Charcot, péninsule antarctique), qui figure en annexe de cette mesure, soit approuvé ; et que
2. Le plan de gestion pour la zone spécialement protégée de l'Antarctique n° 170 annexé à la mesure 16 (2013) soit abrogé.
Annexe
PLAN DE GESTION POUR LA ZONE SPÉCIALEMENT PROTÉGÉE DE L'ANTARCTIQUE NO 170
NUNATAKS MARION, ILE CHARCOT, PÉNINSULE ANTARCTIQUE
Introduction
Afin de protéger les valeurs environnementales et, en particulier, la faune et la flore terrestres des nunataks Marion, sur l'île Charcot dans la péninsule antarctique (69°45' S, 75°15' O), cette zone a été désignée zone spécialement protégée de l'Antarctique (ZSPA).
Le site des nunataks Marion est proche de la côte nord de l'île Charcot, une île éloignée couverte de glace située à l'ouest de l'île Alexandre, dans la partie orientale de la mer de Bellingshausen, le long de la péninsule Antarctique. Il forme une chaîne d'affleurements rocheux longue de 12 km, située au milieu de la côte nord de l'île et s'étend du mont Monique à l'extrémité ouest, jusqu'au mont Martine à l'extrémité est. Sa superficie est de 106,5 km2 (ses dimensions maximales sont de 9,2 km du nord au sud et de 17 km d'est en ouest) et couvre la quasi-totalité, sinon l'intégralité des terres libres de glace de l'île Charcot.
Le peu de visites effectuées par le passé sur cette zone ont rarement duré plus de quelques jours et n'avaient pour but que de mener des recherches géologiques. Toutefois, les scientifiques du British Antarctic Survey (BAS) qui se sont rendus sur place entre 1997 et 2000 ont découvert un site biologique riche situé sur le nunatak de Rils par 69°44'56” S, 75°15'12” O.
Le nunatak de Rils abrite plusieurs caractéristiques exceptionnelles dont deux espèces de lichen qui n'ont été répertoriées nulle part ailleurs en Antarctique, des mousses que l'on trouve rarement sous de latitudes aussi australes et, plus important encore, une absence totale d'arthropodes prédateurs et de collemboles qui sont pourtant présents sur tous les autres sites similaires dans la zone biogéographique. Les nunataks sont extrêmement vulnérables à l'introduction d'espèces non indigènes locales et régionales que des visiteurs pourraient introduire accidentellement sur le site.
La ZSPA n° 170, les nunataks Marion, a été désignée à l'initiative et sur proposition du Royaume-Uni comme ZSPA par la Mesure 4 (2008).
La zone fait partie du système des zones protégées de l'Antarctique, ce qui permet de protéger l'ensemble, unique en son genre, d'espèces trouvées sur les nunataks Marion ; c'est également la première fois que décision est prise de protéger une importante zone de sol représentative de la calotte permanente et des nunataks que l'on trouve couramment dans le sud de la péninsule antarctique. La résolution 3 (2008) recommandait que l'analyse des domaines environnementaux pour le continent antarctique serve de modèle dynamique pour l'identification des zones spécialement protégées de l'Antarctique dans le cadre environnemental et géographique systématisé visé à l'article 3 (2) de l'annexe V du Protocole (voir également Morgan et al., 2007). Dans ce modèle, la ZSPA n° 170 fait partie du Domaine environnemental C (domaine géologique du sud de la péninsule antarctique) et du Domaine E (péninsule antarctique et principaux champs de glaces de l'île Alexandre). D'autres zones protégées contenant le domaine C comprennent la ZSPA n° 147 (bien que non spécifiquement mentionnée dans Morgan et al., 2007). Les autres zones protégées du Domaine E englobent les ZSPA nos 113, 114, 117, 126, 128, 129, 133, 134, 139, 147, 149, 152, et les ZGSA nos 1 et 4. La ZSPA figurant au sein de la Région de conservation biogéographique de l'Antarctique (RCBA) 4, sud central de la péninsule antarctique, est l'une des deux seules ZSPA dans RCBA 4, l'autre étant la ZSPA n° 147 (Terauds et al., 2012).
1. Description des valeurs à protéger
La valeur environnementale exceptionnelle de la zone, qui justifie pleinement sa désignation de ZSPA, repose sur l'existence d'un ensemble d'espèces uniques que l'on peut trouver au sein de son environnement terrestre :
Cette zone se démarque au sein de l'Antarctique maritime par la singularité de sa faune terrestre qui ne semble contenir ni arthropodes prédateurs ni collemboles, contrairement au reste de la zone terrestre environnante où ils demeurent des membres omniprésents et importants. A ce titre, le site offre des possibilités uniques d'étude scientifique des communautés biologiques terrestres de l'Antarctique maritime dont sont absents des éléments écologiques clés.
La flore des nunataks Marion renferme une formation exceptionnelle de trois mousses que l'on trouve rarement au-delà de 65° S (Brachythecum austrosalebrosum, Dicranoweisia crispulaet Polytrichium pilifèrum).
La zone comprend également deux espèces de lichens qui n'avaient jusqu'ici jamais été recensées en Antarctique (Psilolechia lucida et Umbilicaria aff thamnodes) et est la seule région aussi australe à recenser plusieurs espèces de lichens (dont Frutidella caesioatra, Massalongia spp., Ochrolechia. frigida, Usnea aurantiaco-atra et Usnea trachycarpa).
Ces valeurs sont vulnérables aux impacts humains, aux dommages causés par piétinement à l'habitat et à l'introduction d'espèces non indigènes susceptibles de perturber la structure et la fonction de l'écosystème.
2. Buts et objectifs
Les buts et objectifs du plan de gestion sont les suivants :
- éviter la dégradation des valeurs de la zone, ou les préjudices éventuels en empêchant les perturbations humaines injustifiées à la zone ;
- empêcher ou limiter l'introduction de végétaux, animaux et microbes non indigènes dans la zone ;
- réduire les risques d'introduction d'agents pathogènes qui pourraient constituer une source de maladies pour les populations faunistiques de la zone ;
- permettre d'autres travaux de recherche scientifique à condition d'une part, qu'ils résultent de raisons impérieuses qui ne pourraient être satisfaites ailleurs et d'autre part, qu'ils ne portent pas atteinte au système écologique de la zone ; et
- préserver l'écosystème naturel de la zone afin qu'elle serve de zone de référence pour de futures études.
3. Activités de gestion
Les activités de gestion impliquant des visites et l'érection de structures permanentes dans la zone peuvent considérablement accroître les risques d'impact humain irréversibles par l'introduction d'espèces locales non indigènes. Ainsi, afin de gérer au mieux ce site, il convient de mettre l'accent sur la limitation des visites et de l'importation de matériaux au sein de la zone. Les activités de gestion ci-après doivent être entreprises pour protéger les valeurs de la zone :
En raison de la vulnérabilité de la zone et des graves conséquences que pourrait avoir l'introduction d'espèces non indigènes, le nombre des visites de gestion doit être maintenu à son strict minimum et l'érection de structures permanentes, telles que des panneaux d'information et de signalisation sur des sols libres de glace, doit être évitée.
L'autorité nationale doit impérativement informer les équipes de terrain des valeurs qu'il convient de protéger au sein de la zone, ainsi que des précautions et mesures d'atténuation qui sont exposées en détail dans le présent plan de gestion.
Des copies du présent plan de gestion doivent être mises à la disposition des navires et aéronefs prévoyant de visiter les abords de la zone.
Le plan de gestion doit être réexaminé au moins une fois tous les cinq ans et mis à jour en conséquence.
Une copie du présent plan de gestion doit être mise à disposition dans la station de recherche de Rothera (Royaume-Uni ; 67°34' S, 68°07' 0) et de la station General San Martin (Argentine ; 68°68' S, 67°06' 0).
Toutes les activités scientifiques et de gestion entreprises au sein de la zone devraient faire l'objet d'une évaluation d'impact sur l'environnement conformément à ce que requiert l'annexe I du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement.
Les directeurs des programmes antarctiques nationaux en cours d'exécution dans la région se livreront entre eux à des consultations pour veiller à ce que les activités de gestion susmentionnées soient mises en œuvre.
4. Durée de la désignation
La zone est désignée pour une période indéterminée.
5. Cartes
Carte 1. - Situation géographique de l'île Charcot par rapport à l'île Alexandre et à la péninsule antarctique. Spécifications de la carte : WGS84 Stéréographique polaire antarctique. Méridien central : - 55° : Parallèle de référence : - 71°.
Carte 2. - Cartographie de l'île Charcot et matérialisation de la ZSPA n° 170, Nunataks Marion située au nord- ouest de l'île. Spécifications de la carte : WGS84 Stéréographique polaire antarctique. Méridien central : - 75° ; Parallèle de référence : - 71,0°.
Carte 3. - ZSPA n° 170, Nunataks Marion, île Charcot, péninsule antarctique. Spécifications de la carte : WGS84 Stéréographique polaire antarctique. Méridien central : - 75° ; Parallèle de référence : - 71,0° Développée à partir de la mosaïque d'images de l'Antarctique Landsat de l'institut d'études géologiques des Etats-Unis (USGS), ID du lieu : x - 2250000y + 0450000. Métadonnées disponibles sur http://lima.usgs.gov/.
6. Description de la zone
6 (i) Coordonnées géographiques, bornage et caractéristiques du milieu naturel
L'île Charcot, de forme grossièrement circulaire, se situe à environ 50 km de l'île Alexandre dont elle est séparée dans sa partie nord-ouest (à env. 100 km de distance) par le détroit de Wilkins à l'est et le détroit d'Attenborough au sud (cartes 1 et 2). Il y a encore peu de temps, l'île Charcot était reliée à l'île Alexandre par la plateforme glaciaire de Wilkins, mais en 2008, une grande partie s'est effondrée et, en avril 2009, le pont de glace a fini par totalement céder (Vaughan et al., 1993 ; Braun et al., 2009). L'île Charcot est couverte de glace à l'exception des nunataks Marion (69°45' S, 75°15' O) qui forment, au milieu de la côte nord de l'île, une chaîne de 12 km d'affleurements rocheux surplombant la mer, essentiellement constituée de falaises abruptes orientées au nord (Carte 3). Le mont Monique se trouve vers l'extrémité ouest de la chaîne des nunataks Marion et le mont Martine vers son extrémité est. L'altitude de ces deux pics oscille entre 750 et 1 000 mètres.
Les limites de la zone sont établies de la manière suivante :
Le point indiqué sur la côte nord de l'île Charcot par 69°43' 07” S, 75°00'00” O représente l'extrémité nord- ouest de la zone. La limite de la zone s'oriente ensuite à l'ouest en suivant la côte pour parvenir jusqu'au point établi par 69°48'00” S, 75°19'19” O. Puis, elle gagne à l'est et pénètre dans les terres pour atteindre un point de la calotte glaciaire de l'île Charcot par 69°48'00” S, 75°00'00” O. Enfin, elle s'oriente vers le nord pour revenir au point de la côte situé par 69° 43'07” S, 75°00' 00” O. La zone comprend également l'île Cheeseman (69°43'24” S, 75°11'00” O).
Aucune borne ne délimite la zone. Les dimensions maximales sont de 9,2 km du nord au sud et de 17 km d'est en ouest (106,5 km2). La zone comprend également une calotte de glace qui s'étend sur au moins 4 km vers le sud et l'est des nunataks Marion et sert de zone tampon pour empêcher l'introduction accidentelle d'espèces non indigènes (carte 3). Les falaises de glace abruptes de la côte nord de l'île rendent l'accès par la mer difficile.
Conditions climatiques
On ne dispose d'aucune donnée climatique mais l'île Charcot se trouve sur le chemin emprunté par les dépressions qui abordent la péninsule antarctique par l'ouest. Les images satellitaires montrent que la majeure partie du temps, l'île disparaît sous une couverture nuageuse et qu'il faut attendre au mieux la fin de l'été pour qu'elle se libère de la banquise côtière formée au cours de l'hiver, si elle se libère.
Biogéographie
Les recherches réalisées par Smith (1984) et Peat et al., (2007) décrivent les régions biogéographiques établies et reconnues de la péninsule antarctique. L'Antarctique peut être scindée en trois grandes provinces biologiques : maritime nord, maritime sud et continentale. L'île Charcot appartient à la province maritime sud (Smith. 1984), à environ 600 km au nord de la ligne Gressitt, principale discontinuité biogéographique, qui sépare la péninsule antarctique du continent antarctique (Chown et Convey, 2007). Elle relève aussi de la RCBA 4, Sud central de la péninsule antarctique (Terauds et al., 2012).
Géologie
Les roches des nunataks Marion sont constituées de grès et d'argilites turbiditiques similaires à celles trouvées à proximité de l'île Alexandre. Toutefois, la géochronologie et les analyses isotopiques réalisées sur des minéraux détritiques (grains ayant survécu à l'érosion, au déplacement et au dépôt qui conservent les informations relatives à leur roche mère) semblent indiquer que les roches de l'île Charcot diffèrent de celles trouvées sur l'île Alexandre et peut-être aussi de l'ensemble de la péninsule antarctique (Michael Flowerdew, communication personnelle). Les roches de l'île Alexandre se sont formées à partir de sédiments érodés qui se sont détachés de roches présentes sur la péninsule antarctique. Toutefois, les sédiments de l'île Charcot se sont à l'origine déposés à l'intérieur d'une profonde tranchée marine qui est née de la destruction de la plaque Pacifique située en dessous de la bordure de l'ancien continent Gondwana. Des roches sédimentaires ont été arrachées de la plaque Pacifique au cours de sa destruction et se sont accrétées au continent Gondwana, provoquant ainsi leur plissement et leur métamorphose sous l'effet de la haute pression. Les roches sédimentaires de l'île Charcot dateraient du Crétacé (déposées il y a 120 millions d'années environ) et pourraient avoir été transportées sur de longues distances en un laps de temps assez court, avant de se juxtaposer à l'île Alexandre il y a quelques 107 millions d'années.
Biologie
Le site biologique terrestre reconnu (situé sur le nunatak de Rils par 69°44'55” S, 75°15'00” O) s'étend sur approximativement 200 mètres d'est en ouest et sur tout au plus 50 mètres du nord au sud et abrite un vaste biote (Convey et al., 2000). Ce promontoire rocheux végétalisé descend lentement vers le nord-ouest pour ensuite glisser abruptement vers des falaises à pic qui tombent dans la mer. Les visites effectuées sur ce site de décembre 1997 à janvier 2000 ont permis d'observer la présence d'eau libre pendant l'été.
Les biotes de ce site biologique terrestre reconnu sont les suivants :
Bryophytes : 16 mousses (dont Andreaea spp., Bartramia patem, Bryun pseudotriquetrum, Brachytheciun austrosalehrostan, Ceratodon purpureus, Dicranoweisia crispula, Grimmia reflexidens, Hennediella heimii, Hypnwn revolutum, Pohlia spp., Polytrichum piliferum, Schistidium antarctici, Syntrichia princeps) et une hépatique (Cephaloziella varians). Les espèces dominantes sont Andreaea spp., Dicranoweisia crispula et Polytrichum piliferum, que l'on ne rencontre habituellement que dans la région subantarctique. L'abondance de B. austrosalehrosum est d'autant plus remarquable qu'il s'agit d'une espèce hydrique ayant constamment besoin d'eau. L'eau, qui s'écoule au goutte à goutte des couches de neige tardive sur des dalles de roche humide, permet aux mousses de s'implanter pour former des coussins de 15 cm d'épaisseur. (Smith, 1998 ; Convey et al., 2000).
Algue foliacée : Prasiola crispa (Smith, 1998 ; Convey et al. 2000).
Lichens : 34 espèces ont été recensées et deux autres ont pu être assimilées à un genre taxinomique.
Les principales espèces de lichens sont Pseudophebe minuscule, Umbilicaria decussata, Usnea sphacelata et divers taxons encroûtants (Smith, 1998 ; Convey et al., 2000). Des communautés de lichens occupent la plupart des sols et crêtes rocheux, secs et balayés par le vent. Des canaux d'eau de fonte parcourant des dalles rocheuses inclinées sont bordés de grands thalles (pouvant atteindre une quinzaine de centimètres de diamètre) d'Umbilicaria antarctica. La zone dispose de deux espèces de lichens qui n'avaient jamais été recensées en Antarctique jusque-là, (Psilolechia lucida et Umbilicaria aff. thamnodes), mais aussi plusieurs autres espèces de lichens dont la présence, à une latitude si méridionale, est inédite (dont Frutidella caesioatra, Malassalongia spp., Ochrolechia frigida, Usnea aurantiaco-atra et Usnea trachycarpa). Chose inhabituelle, l'espèce très répandue Usnea antarctica n'a pu être observée sur place.
Invertébrés : sept espèces d'acariens, sept nématodes et quatre tardigrades étaient présents dans des prélèvements effectués aux nunataks Marion. Chose surprenante, ni acariens prédateurs ni collemboles n'ont été recensés (Convey, 1999 Convey et al., 2000).
Vertébrés : une petite colonie de soixante manchots Adélie (Pygoscelis adeliae) composée de nombreux oisillons a été aperçue sur les petites îles qui se trouvent juste au nord-ouest du mont Monique (Henderson, 1976 Croxall et Kirkwood, 1979). La colonie a été revue au même endroit en janvier 2011 et se compose de 70 couples reproducteurs et de nombreux oisillons. Il s'agit de la colonie de manchots Adélie la plus au sud de la péninsule antarctique. En dehors de cette colonie de manchots, il n'y a guère de vertébrés dans la zone. Des labbes de McCormick (Stercorarius maccormicki) ont été observés dans la zone mais seul un nid a été découvert sur un tapis de mousse. Parmi les oiseaux observés et susceptibles de se reproduire dans la zone figuraient en petits nombres des sternes antarctiques (Sierna vittata), des pétrels des neiges (Pagodroma nivea), des pétrels antarctiques (Thalassoica antarctica) et des océanites de Wilson (Oceanites oceanicus Kühl) (Henderson, 1976 : Smith, 1998 : Convey et al., 2000).
Bien que tous les éléments du biote recensés soient typiques de la zone biogéographique de l'Antarctique maritime (Smith, 1984), les détails de la composition de leur biome varient considérablement de ceux d'autres communautés répertoriées sur d'autres sites. L'absence apparente de collemboles contraste nettement avec les recensements effectués dans tous les autres sites reconnus de l'Antarctique maritime, où ils sont omniprésents. Plusieurs autres espèces d'animaux aperçues aux nunataks Marion laissent à penser que les densités de population sont comparables à celles trouvées dans de nombreux autres sites côtiers de l'Antarctique maritime et qu'elles ont un ordre de grandeur au moins supérieur à celles habituellement trouvées dans les sites de l'Antarctique continental ou dans la partie sud-est de l'île Alexandre, qui est à la limite sud de l'Antarctique maritime. La contribution numérique des collemboles aux autres faunes de l'Antarctique maritime semble être compensée sur l'île Charcot par la présence de plusieurs acariens prostigmatides de petite taille (Nanorchestes nivalis et Eupodes minutes). L'absence de taxons prédateurs au sein de la communauté d'arthropodes de l'île Charcot est exceptionnelle compte tenu des densités de populations d'arthropodes observées.
Les communautés biologiques terrestres de l'île Charcot sont extrêmement vulnérables à une introduction accidentelle de biotes indigènes et non indigènes de l'Antarctique par l'homme. Convey et al. (2000) écrit :
« Puisque les visiteurs qui débarqueront sur l'île arriveront forcément d'autres sites de la zone maritime [antarctique], ils accroissent le risque de transfert accidentel de terre ou de végétaux agglutinés à leurs chaussures, vêtements, sacs à dos, etc. C'est la raison pour laquelle il est impérieux d'éviter tout transfert d'espèces indigènes entre les diverses populations isolées de l'Antarctique maritime. Il est donc urgent de mettre en place des mesures de contrôle strictes pour tous les visiteurs de ce site, ainsi que d'autres, afin d'assurer la pérennité de leur conservation. ».
Activités humaines antérieures
La zone est extrêmement isolée et très difficile d'accès si ce n'est par aéronef et par conséquent a été visitée par un faible nombre de personnes et ces visites ont généralement été brèves. L'île Charcot a été découverte le 11 janvier 1910 par Jean Baptiste Charcot, alors chef de l'Expédition antarctique française. C'est l'Expédition antarctique de recherches de Ronne (RARE) qui a, la première, atterri sur l'île le 21 novembre 1947 et a profité de l'occasion pour photographier, depuis les airs, des parties de l'île (Searle, 1963).
En novembre 1982, l'Expédition antarctique chilienne et les Forces aériennes chiliennes (FACH) y ont construit une cabane temporaire (30 m2) et une piste d'atterrissage. Le campement était implanté sur la glace à quelques kilomètres à l'est du mont Martine (69°43' S, 75°00' O), à l'endroit même où se situe de nos jours la limite est de la zone. La cabane a été ensevelie par la neige pendant l'hiver 1983 et il ne reste aucune trace de cette station en surface (Comité Nacional de Investigationes Antarcticas (CNIA), 1983 : Veronica Valejos, communication personnelle).
Les géologues et cartographes de la British Antarctic Survey (BAS) ont fait de brèves visites aux nunataks Marion en janvier 1975, du 9 au 13 février 1976, et le 17 janvier 1995. Des biologistes de la BAS ont visité le nunatak de Rils le 22 décembre 1997, les 20 et 21 janvier 1999, le 5 février 1999 et le 16 janvier 2000. Les rapports évaluent à moins de dix le nombre de visites effectuées par les équipes de terrain aux nunataks Marion depuis la première opération menée en 1975. Ces visites n'ont en général pas excédé quelques jours ou quelques heures. Il convient ici de noter que, depuis la découverte de ces écosystèmes uniques en leur genre, plus aucune visite n'a été effectuée aux nunataks Marion à l'intérieur des terres en partant du littoral (Convey et al. 2000). Il est par conséquent probable que l'écosystème ait conservé son état vierge originel et qu'aucune introduction de macrobiote ne se soit produite.
Au début de 2010 et 2011, des scientifiques des États-Unis d'Amérique ont brièvement accosté en bateau pour étudier la colonie de manchots Adélie située sur la côte nord-ouest du mont Monique.
6 (ii) Accès à la zone
Il n'existe pas de points d'accès spécifiques, mais les atterrissages d'aéronefs sont souvent plus sûrs lorsqu'ils sont effectués sur des zones de glace permanente, car accéder à la terre depuis la mer se révèle difficile en raison de la présence de falaises de glace abruptes sur une grande partie du littoral. Les atterrissages d'aéronefs dans la zone doivent être conformes aux conditions décrites dans la section 7 (ii). Au début de 2010 et 2011, de brefs débarquements ont été effectués depuis la mer par des scientifiques américains venus visiter la colonie de manchots Adélie située sur la terre libre de glace au nord-ouest du mont Monique (situation approximative, 69°45'40” S, 75°25'00” O). Les débarquements ont été réalisés en dépit des conditions difficiles, mais habituelles, rencontrées sur la glace de mer. Ces mêmes conditions ont empêché tout débarquement en 2012. C'est pourquoi cet itinéraire n'est pas celui qui est recommandé pour accéder à la zone.
6 (iii) Structures à l'intérieur et à proximité de la zone
Il n'existe aucune installation ou cache connue dans la zone. Un cairn a été construit au sommet du petit nunatak (à environ 126 mètres d'altitude), par 69°44'55” S, 75°15'00” O, à l'occasion du Programme de satellite Doppler mené à l'initiative de l'Institut d'études géologiques des États-Unis (USGS) et de la British Antarctic Survey (BAS) entre 1975 et 1976 (Schoonmaker et Gatson, 1976). Ce cairn de 0,6 cm de hauteur indique, au moyen d'une plaque en cuivre de I'USGS mal rivetée à la roche faillée où est gravée la mention « Jon, 1975- 1976 », l'emplacement de la station Jon. Un poteau de tente en métal (2,4 mètres) a été érigé dans le cairn ; toutefois, il n'en est pas fait mention dans les rapports de visite depuis 1995 (Anonyme, 1977 ; Morgan, 1995).
6 (iv) Emplacement d'autres zones protégées à proximité
Il n'existe pas d'autres ZSPA ou ZGSA à proximité ; la plus proche est la ZSPA n° 147, vallée Ablation dans les monts Ganymède, située à 270 km de la côte est de l'île Alexandre.
6 (v) Zones spéciales au à l'intérieur de la zone
Il n'y a aucune zone spéciale à l'intérieur de la zone.
7. Critères de délivrance des permis
7 (i) Conditions générales pour l'obtention d'un permis
L'accès à la zone est interdit sauf si un permis est délivré par l'autorité nationale compétente conformément au paragraphe 4 de l'article 3 et de l'article 7 de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement.
Les conditions de délivrance d'un permis pour entrer dans la zone sont les suivantes :
- un permis est délivré pour répondre à des missions scientifiques ne pouvant être effectuées ailleurs, ou pour des raisons essentielles à la gestion de la zone ;
- les activités autorisées le sont si, via le processus d'évaluation d'impact sur l'environnement, elles sont considérées comme concourant à la protection continue des valeurs environnementales, écologiques et scientifiques de la zone ;
- les actions permises sont conformes aux objectifs du présent plan de gestion ;
- le permis, ou une copie autorisée, doit être emporté à l'intérieur de la zone ;
- le permis doit être délivré pour une durée déterminée ;
- un rapport est adressé à l'autorité ou aux autorités mentionnées sur le permis ; et
- il convient de notifier à l'autorité compétente toute activité/mesure entreprise qui n'aurait pas été incluse dans le permis accordé.
7 (ii) Accès à la zone et déplacements à l'intérieur de celle-ci
Il est vivement recommandé de se limiter, dans la mesure du possible, à des visites d'une journée sur la zone, afin de réduire les besoins en matériel de campement et les risques connexes d'introduction d'espèces non indigènes dans la zone. S'il n'est pas possible de répondre aux besoins scientifiques ou de gestion en une seule journée, les visites d'une durée plus longue qui exigent de camper à l'intérieur de la zone sont autorisées mais uniquement après que toutes les autres options ont été étudiées en détail et rejetées.
L'accès du personnel ou de matériel arrivant directement d'autres sites de recherche biologique terrestre est interdit dans la zone. Comme le décrit en détail le présent plan de gestion (section 7 [x] ), il est impératif, pour pénétrer dans la zone, que tous les visiteurs et le matériel transitent par une station antarctique ou un navire, où les vêtements et le matériel seront méticuleusement nettoyés.
Pour protéger les valeurs de la zone et limiter le risque d'introduction d'espèces non indigènes, les restrictions mentionnées ci-après s'appliquent à l'intérieur de la zone :
(a) Aéronefs et systèmes d'aéronefs pilotés à distance (RPAS) :
Les aéronefs ne sont autorisés à atterrir dans la zone que s'ils ont pris les mesures décrites en détail dans le présent plan de gestion (section 7 [x] ). Dans le cas contraire, les aéronefs doivent atterrir en dehors de la zone. Au sein de la zone, il est interdit aux aéronefs à voilure fixe et rotative d'atterrir à moins de 100 mètres d'un sol libre de glace et par voie de conséquence, de la flore et de la faune qui lui sont associées. Les 100 mètres restants doivent être parcourus à pied.
Une colonie de manchots Adélie a élu domicile dans la zone, sur la côte située au nord-ouest du mont Monique (situation approximative 69°44'40” S, 75°25'00” O). Les opérations de survol de la zone doivent être réalisées conformément aux « Lignes directrices pour l'exploitation d'aéronefs à proximité des concentrations d'oiseaux dans l'Antarctique », inscrites dans la résolution 2 (2004). Le survol de colonies d'oiseaux dans la zone par des systèmes d'aéronefs pilotés à distance (RPAS) n'est pas autorisé, sauf à des fins scientifiques ou opérationnelles, et en vertu d'un permis émis par une autorité nationale compétente.
(b) Navires et petites embarcations :
Peu d'informations existent sur les zones de débarquement appropriées pour les navires et les petites embarcations (voir section 6 [ii]). Le caractère imprévisible des conditions rencontrées sur la glace de mer dans cette région ne permet pas de recommander le débarquement en bateau comme moyen approprié d'accès à la zone. Toutefois, les débarquements en bateau peuvent être utiles lors de visites sur la côte, pour se rendre par exemple à l'endroit où est implantée la colonie de manchots Adélie au nord-ouest du mont Monique (situation approximative 69°45'40” S, 75°25'00” O).
(c) Véhicules terrestres et traîneaux :
Aucun véhicule terrestre ne doit être introduit dans la zone, sauf pour des raisons scientifiques, de gestion ou de sécurité indispensables. Les véhicules terrestres et traîneaux ne sont autorisés dans la zone que s'ils ont pris les mesures décrites en détail dans le présent plan de gestion (section 7 [x] ). Au sein de la zone, il est interdit aux scooters des neiges, traîneaux et tout autre véhicule terrestre de s'approcher à moins de 100 mètres d'un sol libre de glace et par voie de conséquence, de la flore et de la faune qui lui sont associées. Les 100 mètres restants doivent être parcourus à pied.
(d) Déplacements humains :
Le trafic piétonnier sera maintenu au strict minimum pour garantir le bon déroulement des activités autorisées. Lorsqu'aucune voie piétonnière n'est identifiée, les déplacements piétonniers devraient être strictement réservés à la réalisation d'activités autorisées et toutes les mesures nécessaires devraient être prises pour éviter les effets de piétinement. Les visiteurs doivent éviter de marcher sur des aires végétalisées et progresser prudemment sur les sols humides, en particulier les lits des cours d'eau pour préserver de leurs pas les sols fragiles, les communautés végétales et algales ainsi que la qualité de l'eau.
Des mesures de quarantaine rigoureuses doivent être prises à titre de précaution, tel que cela est décrit à la section 7 (x) du présent plan de gestion.
7 (iii) Activités pouvant être conduites à l'intérieur de la zone
Les activités pouvant être conduites sont :
- les travaux de recherche scientifiques impérieux qui ne peuvent être réalisés ailleurs et ne mettront pas en danger l'écosystème de la zone ;
- les activités d'échantillonnage nécessaires pour que les programmes de recherches soient validés ; et
- les activités indispensables de gestion et de surveillance.
7 (iv) Installation, modification ou enlèvement de structures
Aucune structure ou équipement scientifique ne doit être érigé ou installé dans la zone, hormis pour satisfaire des raisons scientifiques ou de gestion impérieuses ; dans ce cas, un permis précisant la durée de ces activités doit être délivré.
Les structures ou installations permanentes sont interdites.
Toutes les bornes, les structures et tout l'équipement scientifique installés dans la zone doivent être clairement identifiés et mentionner le pays, le nom du principal chercheur ou de l'agence, l'année d'installation et la date prévue d'enlèvement.
Tous ces articles doivent être exempts d'organismes, de propagules (par ex. semences, œufs et spores) et de terre non stérile (voir section 7 [x] ) ; ils doivent être constitués de matériaux pouvant résister aux conditions environnementales et présenter un risque marginal de contamination de la zone.
L'enlèvement de structures et équipements spécifiques dont le permis a expiré incombe à l'autorité qui a, en premier lieu, délivré le permis et doit en outre être l'un des critères de délivrance du permis.
Les structures existantes ne peuvent être démantelées sans délivrance de permis.
7 (v) Emplacement des camps
Les campements dans la zone ne sont autorisés que si les objectifs scientifiques ou de gestion requièrent plus d'une journée pour être menés à bien. Le campement au sein de la zone est également autorisé dans des situations d'urgence. A moins que cela ne se révèle inévitable pour des raisons de sécurité, les tentes doivent être installées sur de la neige ou de la glace, à 500 mètres au moins de la première zone libre de glace.
L'équipement nécessaire à l'établissement de campements au sein de la zone doit être nettoyé et transporté comme cela est décrit à la section 7 (x) du présent plan de gestion.
7 (vi) Restrictions relatives aux matériaux et organismes pouvant être introduits dans la zone
Outre les exigences du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement, les restrictions concernant l'apport de matériaux ou d'organismes au sein de la zone sont les suivantes :
L'introduction délibérée d'animaux, de végétaux, de micro-organismes et de terre non stérile dans la zone est interdite.
Il est nécessaire de prendre des précautions pour éviter l'introduction accidentelle d'animaux, de végétaux, de micro-organismes et de terre non stérile provenant d'autres régions biologiquement distinctes (au sein ou en dehors de la zone couverte par le Traité sur l'Antarctique). Les visiteurs doivent également consulter et se conformer aux recommandations appropriées figurant dans le Manuel sur les espèces non indigènes du Comité pour la protection de l'environnement (CPE, 2011) et dans l'Environmental code of conduct for terrestrial scientific field research in Antarctica [Code de conduite environnemental pour la recherche scientifique terrestre de terrain dans l'Antarctique] (SCAR, 2009). Des mesures supplémentaires relatives à la biosécurité spécifiques à ce site sont énumérées à la section 7 (x).
Aucun produit avicole, y compris les produits alimentaires contenant de la poudre d'œufs non cuits, ne doit être introduit dans la zone.
Aucun herbicide ou pesticide ne doit être introduit dans la zone. Les autres produits chimiques, y compris les radionucléides ou les isotopes stables pouvant être introduits pour répondre à un besoin scientifique impérieux et autorisés par un permis, doivent être enlevés de la zone avant ou au moment de la conclusion de l'activité qui a donné lieu à la délivrance du permis. Les émissions irrémédiables de radionucléides ou d'isotopes stables dans l'environnement doivent impérativement être évitées.
Le combustible, les aliments et autres matériaux ne peuvent être entreposés dans la zone, sauf en cas de nécessité absolue liée aux activités pour lesquelles le permis a été délivré. Ils doivent être stockés et manipulés de façon à limiter le risque d'introduction accidentelle dans l'environnement. Le combustible, les aliments et autres matériaux doivent être uniquement entreposés sur des zones de neige ou de glace et éloignés d'au moins 500 mètres du sol libre de glace le plus proche. Les dépôts permanents sont interdits.
Les matériaux introduits dans la zone le sont pour une période donnée uniquement et doivent être enlevés lors de, ou avant la conclusion de ladite période.
7 (vii) Prélèvement de végétaux et capture d'animaux ou perturbations nuisibles à la faune et la flore
Le prélèvement de végétaux ou toutes perturbations nuisibles à la faune et la flore sont interdits, sauf s'ils sont, d'une part, autorisés par un permis et d'autre part, conformes à l'annexe II du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement. Tous prélèvements ou toutes perturbations nuisibles aux animaux, doivent au minimum se faire conformément aux dispositions du SCAR code of conduct for the use of animals for scientific purposes in Antarctica [Code de conduite du SCAR pour l'utilisation d'animaux à des fins scientifiques dans l'Antarctique] (2011). Les activités d'échantillonnage du sol ou de la végétation doivent être limitées au strict minimum requis à des fins scientifiques ou à des fins de gestion et exécutées avec des techniques qui minimisent les perturbations susceptibles d'être causées au sol, aux structures de glace et au biote.
7 (viii) Collecte ou retrait de matériaux non introduits dans la zone par le titulaire du permis
Le ramassage ou l'enlèvement de matériaux dans la zone ne peut se faire que sur délivrance d'un permis et doit se limiter au minimum requis pour les activités menées à des fins scientifiques ou de gestion. Tous matériaux d'origine anthropique susceptibles d'avoir un impact sur les valeurs de la zone et n'ayant pas été introduits par le titulaire du permis ou une autre personne autorisée peuvent être enlevés dans la mesure où cet enlèvement n'entraînera pas de conséquences plus graves que de les laisser in situ. Dans ce cas, les autorités compétentes doivent en être informées et donner leur accord.
7 (ix) Elimination des déchets
Tous les déchets, y compris les déchets humains, doivent être retirés de la zone.
7 (x) Mesures qui peuvent être nécessaires pour continuer de répondre aux objectifs du plan de gestion
Pour concourir à la protection des valeurs écologiques et scientifiques de la zone, il est nécessaire d'en garantir l'isolement et de maintenir un risque faible d'impact humain. Les visiteurs doivent donc prendre des mesures de précaution particulières pour limiter l'introduction d'espèces non indigènes. D'autres conseils sont donnés dans le Manuel sur les espèces non indigènes du CPE (édition 2017) et l'Environmental code of conduct for terrestrial scientific field research in Antarctica [Code de conduite environnementale pour la recherche scientifique sur le terrain en Antarctique] (SCAR 2009). Une vigilance particulière doit être accordée à l'introduction d'animaux et de végétaux issus de :
- sols provenant d'autres sites de l'Antarctique, y compris ceux qui sont proches des stations ;
- sols en provenance de régions extérieures à l'Antarctique.
Pour entrer dans la zone, les visiteurs doivent limiter les risques d'introduction en se conformant aux mesures suivantes :
(a) Aéronefs :
L'inspection minutieuse ainsi que le nettoyage intérieur et extérieur des aéronefs doivent être réalisés juste avant leur envol de la station antarctique ou du navire. L'intérieur des aéronefs doit être méticuleusement balayé et aspiré, tandis que l'extérieur doit être brossé ou nettoyé à la vapeur. Tous les aéronefs ayant, depuis leur nettoyage à la station ou sur le navire, atterri sur d'autres pistes rocheuses ou à proximité de sites biologiquement riches sont interdits d'accès à la zone.
Avant d'atterrir dans la zone, les aéronefs à voilure fixe ayant décollé d'une piste en gravier doivent avoir atterri ou posé leurs skis sur de la neige propre en dehors de la zone afin d'en déloger toute la terre qui aurait pu y adhérer.
(b) Petites embarcations :
Les embarcations servant à transporter les visiteurs depuis un navire de soutien jusqu'à la limite de la zone doivent être nettoyées (surtout à l'intérieur) et être exemptes de terre, de saletés et de propagules.
(c) Véhicules terrestres et traîneaux :
Toutes traces excessives de boue, de terre, de végétaux et de saleté doivent être ôtées des véhicules terrestres et des traineaux pénétrant dans la zone. Cette opération doit être idéalement effectuée avant que les véhicules ne quittent la station ou le navire qui les transférait sur le terrain. Les véhicules terrestres propres mais ayant roulé depuis dans des zones rocheuses ou terreuses ne doivent pas pénétrer dans la zone.
(d) Equipements nécessaires à l'établissement d'un camp :
Tous les équipements de camping, y compris l'équipement d'urgence, doivent être minutieusement nettoyés (débarrassés de terre et de propagules et, dans la mesure du possible, placés dans des sacs ou des bâches en plastique) avant d'être introduits dans la zone. Cela inclut également le matériel de camping d'urgence transporté par un aéronef qui atterrit dans la zone.
(e) Matériel d'échantillonnage, appareils scientifiques et balises de terrain :
Tout le matériel d'échantillonnage, tous les appareils scientifiques et toutes les balises introduits dans la zone doivent avoir été stérilisés et maintenus en cet état avant d'être utilisés à l'intérieur de la zone. La stérilisation doit se faire au moyen d'une méthode convenue, que ce soit au moyen de rayons UV, d'un autoclavage ou par la stérilisation de la surface en utilisant 70 % d'éthanol ou un biocide disponible dans le commerce (du Virkon® par exemple) (se reporter au « Environmental code of conduct for terrestrial scientific field research in Antarctica » [Code de conduite environnemental pour la recherche scientifique terrestre de terrain dans l'Antarctique] » [SCAR, 2009] ).
(f) Matériel d'usage général :
Le matériel général comprend les harnais, les crampons, le matériel d'escalade, les piolets, les bâtons de marche, le matériel de ski, les balises temporaires, les luges, les traîneaux, les appareils photographiques et vidéo, les sacs à dos, les boîtes de traîneaux ainsi que tout autre équipement personnel.
Tout le matériel destiné à être utilisé à l'intérieur de la zone doit être dépourvu de propagules biologiques comme les semences, les œufs, les insectes, les fragments de végétaux et la terre. Tout le matériel utilisé ou introduit dans la zone doit, dans la mesure du possible, avoir été nettoyé consciencieusement et stérilisé à la station antarctique ou sur le navire d'où ils proviennent. Le matériel doit avoir été maintenu dans cet état avant d'entrer dans la zone et être idéalement scellé dans des sacs en plastique ou autres conteneurs propres.
(g) Vêtements de dessus :
Les vêtements de dessus comprennent les chapeaux, les gants, les pulls ou les polaires, les vestes, les pantalons en toile ou en laine, les pantalons ou les salopettes imperméables, les chaussettes, les chaussures ainsi que tout autre vêtement susceptible d'être exposé à l'environnement. Tout le matériel destiné à être utilisé à l'intérieur de la zone doit être dépourvu de propagules biologiques comme les semences, les œufs, les insectes, les fragments de végétaux et la terre. Les chaussures et vêtements de dessus utilisés ou introduits dans la zone doivent, dans la mesure du possible, avoir été préalablement lavés en machine. Les semences et propagules prisonnières des bandes Velcro® doivent être minutieusement ôtées. Les vêtements neufs, directement sortis de l'emballage du fabricant juste avant d'entrer dans la zone, n'ont pas à être nettoyés.
Les procédures additionnelles pour s'assurer que des espèces non indigènes ne soient pas introduites dans la zone sur des chaussures et des vêtements dépendent de la façon dont le visiteur est entré sur la zone :
(i) en atterrissant directement dans la zone ; (ii) par voie terrestre depuis l'extérieur vers l'intérieur de la zone ou (iii), par bateau jusqu'aux limites de la zone :
i. Atterrissage directement au sein de la zone. Une surcombinaison de protection stérile doit être portée. Ce vêtement de protection doit être enfilé juste avant de quitter l'aéronef. Les chaussures de rechange, nettoyées auparavant à l'aide d'un biocide puis scellées dans des sacs en plastique, doivent être déballées et chaussées juste avant d'entrer dans la zone.
ii. Déplacement par voie terrestre depuis l'extérieur vers l'intérieur de la zone.
Il n'est pas recommandé de porter une surcombinaison de protection stérile car, une fois à l'intérieur de la zone, beaucoup de déplacements sur un sol crevassé sont susceptibles d'avoir lieu et l'emploi d'une telle surcombinaison peut limiter l'utilisation de matériel de sécurité comme des cordes et des harnais. Les déplacements par voie terrestre au sein de la zone peuvent faire l'objet de mesures alternatives. Chaque visiteur est tenu d'apporter au moins deux jeux de vêtements de dessus. Le premier jeu sera porté pour effectuer le voyage jusqu'à la limite de la zone. Le second, qui a déjà été nettoyé et scellé dans des sacs en plastique, ne doit être porté qu'à l'intérieur de la zone. Juste avant de pénétrer dans la zone, les visiteurs doivent changer de vêtements pour porter le jeu propre. Les chaussures de rechange, nettoyées auparavant à l'aide d'un biocide puis scellées dans des sacs en plastique, doivent être déballées et chaussées juste avant d'entrer dans la zone. Les vêtements de dessus sales qui ont été enlevés doivent être conservés dans des sacs en plastique étiquetés et scellés, de préférence en dehors de la zone. Lorsque les visiteurs quittent la zone par voie terrestre, les vêtements portés dans la zone doivent être enlevés et placés dans un sac en plastique propre et étiqueté et soit, resservir lors de visites ultérieures de la zone, soit renvoyés pour nettoyage à la station antarctique ou au navire d'origine.
iii. Déplacement par bateau jusqu'aux limites de la zone : chaque visiteur, y compris l'équipage embarqué, doit, alors qu'il est encore à bord du bateau d'assistance, et juste avant d'embarquer sur la petite embarcation devant leur permettre d'atteindre la zone, enfiler des vêtements propres (vêtements de navigation, gilets de survie et chaussures) dépourvus de terre, de semences ou de toutes autres propagules.
S'ils le préfèrent, les visiteurs peuvent, lors de leur arrivée à la limite de la zone et avant de quitter le bateau, revêtir des surcombinaisons de protection. Les autres vêtements et chaussures nécessaires pour permettre l'entrée des visiteurs au sein de la zone, doivent être nettoyés avant de quitter le bateau d'assistance et demeurer stockés dans un contenant scellé (sac en plastique) jusqu'à leur utilisation.
7 (xi) Rapports de visite
Le principal titulaire de chaque permis délivré soumet dès que possible à l'autorité nationale compétente, et dans un délai de maximum de six mois après la fin de la visite, un rapport décrivant les activités menées dans la zone lors de chaque visite réalisée. Ces rapports doivent inclure les renseignements identifiés dans le formulaire du rapport de visite d'une zone spécialement protégée de l'Antarctique inclus dans le Guide pour la préparation des plans de gestion de zones spécialement protégées en Antarctique (annexe 2). Ce rapport doit précisément mentionner les zones libres de glace spécifiques visitées au sein de la zone (en indiquant, dans la mesure du possible, les coordonnées GPS de ces sites), le temps passé à chaque endroit et les activités menées. L'autorité nationale doit, chaque fois qu'elle le peut, également transmettre une copie du rapport de visite à la Partie étant à l'initiative du plan de gestion, afin de l'aider à la gestion de la zone et dans la révision du plan de gestion. Les Parties doivent, dans la mesure du possible, déposer les originaux ou les copies de ces rapports dans une archive à laquelle le public pourra avoir accès afin de maintenir ainsi une archive d'usage. Cette archive pourra être utilisée à la fois lors de tout réexamen du plan de gestion et dans le cadre de l'organisation de l'utilisation scientifique du site.
8. Bibliographie
Anonyme (1977). British Antarctic Survey Archives Service, Arc. Ref. ES2/EW360.1/SR17-18/7,8. Réunion consultative du Traité sur l'Antarctique (2004). Lignes directrices pour les aéronefs à proximité des concentrations d'oiseaux en Antarctique. résolution 2 de la RTCA (2004) :
Braun, M. , Humbert, A., and Moll, A. (2009). Changes of Wilkins Ice Shelf over the past 15 years and inferences on its stability. The Cryosphere 3 : 41-56.
Comite Nacional de Investigaciones Antarcticas. (1983). Informe de las actividades Antarcticas de Chile al SCAR.Santiago, Instituto Antarctico Chileno. Santiago, Instituto Antarctico Chileno.
Comité pour la protection de l'environnement (CPE) (2017). Manuel sur les espèces non indigènes - 2e édition. Manuel préparé par le Groupe de contact intersessions du CPE et adopté par la Réunion Consultative du Traité sur l'Antarctique par la résolution 4 (2016). Buenos Aires, Secrétariat du Traité sur l'Antarctique.
Chown, S. L., and Convey, P. (2007). Spatial and temporal variability across life's hierarchies in the terrestrial Antarctic. [Variabilité dans les hiérarchies vivantes dans les terres antarctiques] Philosophical Transactions of the Royal Society B 362 (1488) : 2307-2R31.
Convey, P. (1999). Terrestrial invertebrate ecology. Rapport interne non publié de la British Antarctic Survey Ref. R/1998/NT5.
Convey, P., Smith, R. I. L., Peat, H. J. and Pugh, P. J. A. (2000). The terrestrial biota of Charcot Island, eastern Bellingshausen Sea, Antarctica : an example of extreme isolation. Antarctic Science 12 : 406-413.
Croxall, J. P., and Kirkwood, E. D. (1979). The distribution of penguins on the Antarctic Peninsula and islands of the Scotia Sea. British Antarctic Survey, Cambridge.
Henderson, I. (1976). Summer log of travel and work of sledge kilo in northern Alexander Island and Charcot Island, 1975/1976. Rapport interne non publié de la British Antarctic Survey Ref. T/1975/K11.
Morgan, F., Barker, G., Briggs, C., Price, R., and Keys, H. (2007). Environmental Domains of Antarctica Version 2.0 Final Report. Landcare Research Contract Report LC0708/055.
Morgan, T. (1995). Sledge echo travel report, 1994/5 season - geology in central Alexander Island. Rapport interne non publié de la British Antarctic Survey Ref. R/1994/K7.
Peat, H. J., Clarke, A., et Convey, P. (2007). Diversity and biogeography of the Antarctic flora. Journal of Biogeography 34 : 132-146.
Schoonmaker, J. W., et Gatson, K. W. (1976). U. S. Geological Survey/British Antarctic Survey Landsat Georeceiver Project. British Antarctic Survey Archives Service, Arc. Ref. ES2/EW360/56.
SCAR (Comité scientifique pour la recherche antarctique) (2009). Environmental code of conduct for terrestrial scientific field research in Antarctica [Code de conduite environnementale pour la recherche scientifique sur le terrain en Antarctique]. XXXIIe RCTA IP4.
SCAR (Comité scientifique pour la recherche antarctique) (2011). SCAR code of conduct for the use of animals for scientific purposes in Antarctica [Code de conduite du SCAR pour l'utilisation d'animaux à des fins scientifiques]. XXXIIe RCTA IP53.
Searle, D. J. H. (1963). The evolution of the map of Alexander and Charcot Islands, Antarctica. The Geographical Journal 129 : 156-166.
Smith, R. I. L. (1984). Terrestrial plant biology of the sub-Antarctic and Antarctic. Dans : Antarctic Ecology, Vol. 1. Rédacteur : R. M. Laws. London, Academic Press.
Smith, R. I. L. (1998). Field report : sledge delta, November 1997 - January 1998. Rapport interne non publié de la British Antarctic Survey Ref. R/1997/NT3.
Terauds, A., and Lee, J. R. (2016). Antarctic biogeography revisited : updating the Antarctic Conservation Biogeographic Regions. Diversity and Distribution 22 : 836-840.
Terauds, A., Chown, S. L., Morgan, F., Peat, H. J., Watt, D., Keys, H., Convey, P., and Bergstrom, D. M. (2012). Conservation biogeography of the Antarctic. Diversity and Distributions 18 : 726-41.
Vaughan, D. G., Mantripp, D. R., Sievers, J., and Doake C. S. M. (1993). A synthesis of remote sensing data on Wilkins Ice Shelf, Antarctica. Annals of Glaciology : 17 : 211-218.
Carte 1. - Situation géographique de l'île Charcot par rapport à l'île Alexandre et à la péninsule antarctique. Spécifications de la carte : WGS84 Stéréographique polaire antarctique. Méridien central : - 55° : Parallèle de référence : - 71°.
Carte 2. - Cartographie de l'île Charcot et matérialisation de la ZSPA n° 170, Nunataks Marion située au nord- ouest de l'île. Spécifications de la carte : WGS 1984 Stéréographique polaire antarctique. Méridien central : - 75° ; Parallèle de référence 1 : - 71,0°.
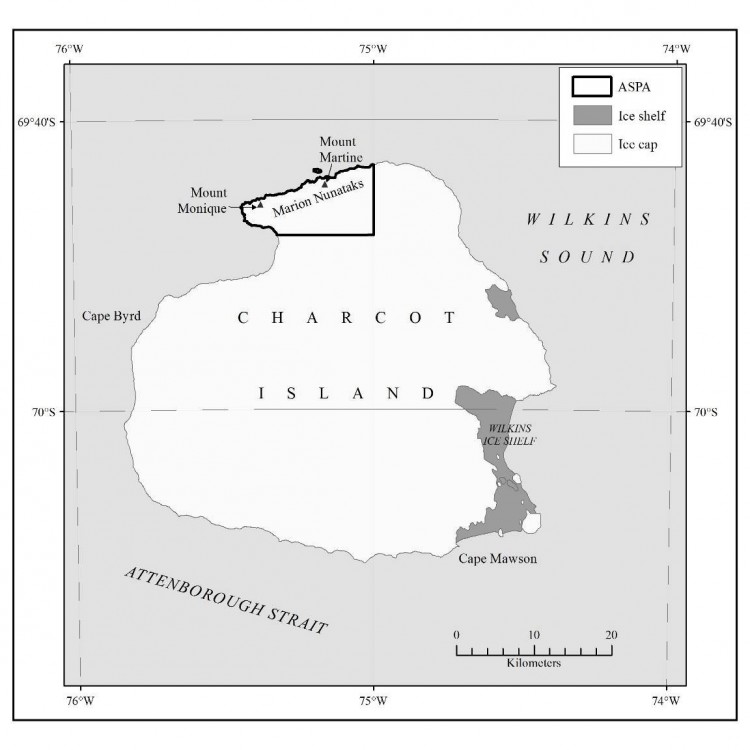
Carte 3. - ZSPA n° 170, Nunataks Marion, île Charcot, péninsule antarctique. Spécifications de la carte : WGS 1984 Stéréographique polaire antarctique. Méridien central : - 75'S ; Parallèle de référence 1 : - 71,0°. Développée à partir de la mosaïque d'images de l'Antarctique Landsat de l'Institut d'études géologiques des Etats-Unis (USGS), ID du lieu : x - 2250000y + 0450000. Métadonnées disponibles sur http://lima.usgs.gov/.
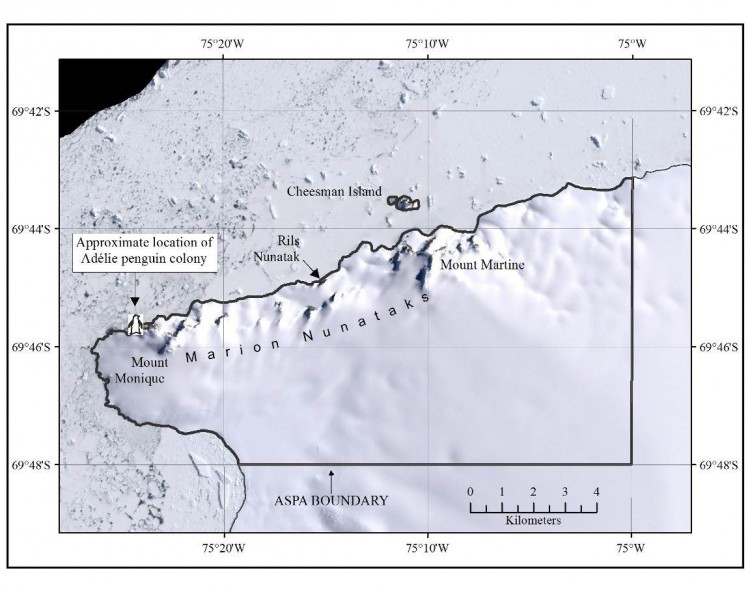
Annexe
MESURE 6 (2018)
ZONE SPÉCIALEMENT PROTÉGÉE DE L'ANTARCTIQUE NO 172 (PARTIE INFÉRIEURE DU GLACIER TAYLOR ET BLOOD FALLS DE LA VALLÉE TAYLOR, DANS LES VALLÉES SÈCHES DE MCMURDO EN TERRE VICTORIA) : PLAN DE GESTION RÉVISÉ (ENSEMBLE UNE ANNEXE)
Les Représentants,
Rappelant les articles 3, 5 et 6 de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement qui disposent de la désignation des zones spécialement protégées de l'Antarctique (« ZSPA ») et de l'adoption des plans de gestion pour ces zones ;
Rappelant la mesure 9 (2012) qui a désigné la partie inférieure du glacier et les Blood Falls de la vallée Taylor, vallées sèches de McMurdo, terre Victoria comme la ZSPA n° 172 et a adopté un plan de gestion pour la zone ;
Notant que le Comité pour la protection de l'environnement a approuvé un plan de gestion révisé pour la ZSPA n° 172 ;
Souhaitant remplacer le plan de gestion actuel pour la ZSPA n° 172 avec le plan de gestion révisé ;
Recommandent à leurs Gouvernements d'approuver la mesure suivante conformément au paragraphe 1 de l'article 6 de l'annexe V du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement :
Que :
1. Le plan de gestion révisé pour la zone spécialement protégée de l'Antarctique n° 172 (Partie inférieure du glacier et Blood Falls de la vallée Taylor, vallées sèches de McMurdo, terre Victoria), qui figure en annexe de cette mesure, soit approuvé ; et
2. Le plan de gestion pour la zone spécialement protégée de l'Antarctique n° 172 annexé à la mesure 9 (2012) soit abrogé.
Annexe
PLAN DE GESTION POUR LA ZONE SPÉCIALEMENT PROTÉGÉE DE L'ANTARCTIQUE N° 172
INFÉRIEURE DU GLACIER TAYLOR ET BLOOD FALLS DE LA VALLÉE TAYLOR, VALLÉES SÈCHES DE McMURDO, TERRE VICTORIA
Introduction
Les Blood Falls sont une coulée d'eau saline riche en fer située à l'extrémité du glacier Taylor, dans la vallée Taylor, vallées sèches de McMurdo. La coulée semble être un vaste aquifère de saumure sous-glaciaire sous la longueur mesurable (env. 5 km) de la zone d'ablation du glacier Taylor, située à une distance estimée entre 1 et 6 km en amont des Blood Falls. Emplacement approximatif et coordonnées : zone en sous-surface 436 km2 (coordonnées centrales 161°40,230'E, 77°50,220'S) ; zone subaérienne 0,11 km2 (centrée sur la coulée des Blood Falls à 162°15,809'E, 77°43.365'). Les raisons principales pour lesquelles cette zone a été désignée sont ses caractéristiques physiques uniques, ainsi que son écologie microbienne et sa géochimie inhabituelles. Cette zone est un site important pour la recherche exobiologique et représente une opportunité unique de prélever des échantillons de l'environnement sous-glaciaire sans contact direct. L'influence des Blood Falls sur le lac Bonney avoisinant présente aussi un grand intérêt scientifique. Par ailleurs, la zone d'ablation du glacier Taylor est un important site de recherche paléoclimatique et glaciologique. Le réservoir sous-glaciaire d'eau saumâtre de la partie inférieure du glacier Taylor et les Blood Falls sont uniques au monde et constituent un site d'intérêt scientifique exceptionnel. La désignation de cette zone permet aux chercheurs d'étudier la glace située au cœur du glacier Taylor, à condition que des mesures soient mises en place afin d'éviter que ces activités ne nuisent au réservoir et au système hydrologique des Blood Falls. Selon l'Analyse des domaines environnementaux du continent antarctique (résolution 3 [2008] ), cette zone se situe dans l'environnement S - Géologique de McMurdo - Terre Victoria du Sud. D'après la classification des régions de conservation biogéographiques de l'Antarctique (v2) (résolution 3 [2017] ), la zone se trouve dans la RCBA 9 - Terre Victoria du Sud.
1. Description des valeurs à protéger
Les Blood Falls sont une caractéristique particulière du glacier situées à 162°16,288'E, 77°43,329'S, à l'extrémité du glacier Taylor dans la vallée Taylor, vallées sèches de McMurdo, terre Victoria du Sud (carte 1).
Cet élément unique se forme à l'endroit où un débit d'eau saline d'origine sous-glaciaire, riche en fer, émerge à la surface et s'oxyde rapidement, acquérant de ce fait une couleur rouge très particulière (image 1). Les données disponibles indiquent que la coulée provient d'un réservoir sous-glaciaire contenant des dépôts de sel marin et de l'eau saumâtre, situé sous le glacier Taylor (Keys 1980 ; Hubbard et al. 2004 ; Mikucki et al. 2015) (carte 1). C'est une caractéristique unique de par sa configuration physique, sa biologie microbienne et sa géochimie, qui a une influence importante sur l'écosystème local du lac Bonney. Les épisodes de débit des Blood Falls fournissent par ailleurs une occasion unique d'échantillonner les propriétés du réservoir sous-glaciaire et de son écosystème.
C'est en 1911 que Griffith Taylor, le géologue principal de Robert F. Scott, avait observé pour la première fois les Blood Falls. Ses caractéristiques morphologiques et géochimiques inhabituelles n'ont toutefois pas commencé à être étudiées avant la fin des années 1950 (Hamilton et al. 1962 ; Angino et al. 1964 ; Black et al. 1965). La caractéristique dénommée Blood Falls est le principal site de débit du front glaciaire du glacier Taylor (carte 2). On a aussi observé un débit d'eau saline latérale secondaire à la surface de sédiments, à environ 40 mètres au nord du glacier Taylor, au bord du delta du cours d'eau Santa Fe (162° 16,042'E, 77°43,297'S, carte 2). L'emplacement et la forme exacts du réservoir sous-glaciaire alimentant les Blood Falls ne sont pas encore connus, mais les résultats des travaux de cartographie géologique, glacio-chimique et géophysique indiquent que le réservoir s'étend sous le lac Bonney et le front glaciaire sur une distance d'au moins 5 km (Keys 1980 ; Hubbard et al. 2004 ; Mikucki et al. 2015, Foley et al. 2015). On estime que le réservoir d'eau saumâtre a été enserré par la glace il y a environ 3 à 5 Ma BP (Marchant et al. 1993) et qu'il représente peut-être la caractéristique liquide la plus ancienne de la vallée Taylor (Lyons et al. 2005).
La coulée des Blood Falls contient une communauté microbienne unique, qui semble d'origine marine. Les microbes qu'on y trouve peuvent survivre dans un environnement sous-glaciaire pendant des millions d'années sans aucun apport en carbone venant de l'extérieur. La haute teneur en fer et en sel de l'écosystème microbien des Blood Falls, ainsi que son emplacement sous la glace du glacier, en fait un site important pour les études exobiologiques, et pourrait fournir un analogue des conditions pouvant apparaître sous les calottes de glace polaires de Mars ou dans les mondes-océans comme Encelade ou Europe. Il est donc essentiel de protéger la communauté microbienne des Blood Falls, le réservoir d'eau saumâtre et le système hydrologique sous-glaciaire qui lui est associé.
Le débit épisodique des Blood Falls dans le lac Bonney adjacent transforme la composition géochimique du lac et lui fournit des nutriments qui y sont autrement limités, faisant de ce site un site important pour étudier l'impact des écoulements sous-glaciaires sur les écosystèmes lacustres. De plus en plus d'indices montrent que la saumure de l'aquifère sous-glaciaire est directement connectée à l'eau du fond du lac Bonney dans la zone sous-glaciaire (Mikucki et al. 2015 ; Spigel et al. sous presse 2018).
Le glacier Taylor est un site important pour les études glaciologiques et paléoclimatiques en Antarctique. Il offre une occasion unique d'étudier la décharge des glaciers antarctiques en fonction des changements environnementaux, à partir des données paléoclimatiques des carottes de glace du dôme Taylor, des données géologiques de la vallée Taylor et des données climatiques des sites avoisinants du programme US Long Term Ecological Research (LTER) (Kavanaugh et al. 2009 a ; Bliss et al. 2011). La zone d'ablation inférieure du glacier Taylor a été identifiée comme étant un site potentiellement important pour les études paléoclimatiques car elle expose de la glace formée lors de la dernière période glaciaire et permet de mesurer avec une résolution temporelle élevée les concentrations antérieures de gaz en traces (Aciego et al. 2007). Le glacier Taylor a aussi une valeur scientifique importante pour les études glaciologiques, en particulier la dynamique des glaciers et la relation entre les contraintes et le flux glaciaire, ainsi que pour d'autres études glaciologiques (Kavanaugh & Cuffey 2009).
Le système des Blood Falls est un site de grande valeur pour la recherche microbiologique, hydrochimique, glaciologique et paléoclimatologique. Les aspects les plus étonnants du système des Blood Falls sont sa configuration physique, la chimie de son eau saumâtre et son écosystème microbien. Les Blood Falls ont aussi un impact considérable sur la géochimie et la microbiologie du lac Bonney. La zone possède des valeurs esthétiques exceptionnelles et une valeur pédagogique importante, ce site ayant fait l'objet de toute une série d'articles scientifiques et médiatiques au cours des dernières années. Les Blood Falls et le réservoir d'eau saumâtre du glacier Taylor méritent une protection spéciale en raison de leurs valeurs scientifiques exceptionnelles, de leur configuration unique, de leur origine ancienne, de leur importance pour les écosystèmes de la zone environnante et de leur vulnérabilité aux activités humaines.
D'après les connaissances actuelles, le mécanisme de contamination éventuelle du réservoir d'eau saumâtre du glacier Taylor le plus probable serait une contamination directe du réservoir sous-glaciaire ou bien de zones du substrat, qui pourrait par la suite libérer des fluides sous-glaciaires dans le réservoir. Mais il est difficile d'évaluer la probabilité de ce mécanisme en raison des incertitudes relatives à l'emplacement du réservoir sous-glaciaire et à la façon dont il est relié au système hydrologique sous-glaciaire. C'est pourquoi le principe de précaution est appliqué quand il s'agit de définir les limites de la composante en sous-surface de la zone.
2. Buts et objectifs
La gestion de la partie inférieure du glacier Taylor et des Blood Falls vise à :
- éviter toute détérioration ou tout risque de détérioration des valeurs de la zone en empêchant toute perturbation humaine ou tout échantillonnage inutile dans la zone ;
- permettre la réalisation dans la zone de travaux de recherche scientifique, en particulier sur la communauté microbienne, la chimie de l'eau et la configuration physique de la partie inférieure du glacier Taylor et des Blood Falls ;
- permettre d'autres travaux de recherche scientifique et des visites pédagogiques ou de vulgarisation scientifique qui ne portent pas atteinte aux valeurs de la zone ;
- réduire au minimum l'éventualité d'introduire des plantes, des animaux et des microbes non indigènes dans la zone ; et
- permettre des visites à des fins de gestion conformément aux objectifs du plan de gestion.
3. Activités de gestion
Les activités de gestion ci-dessous seront menées à bien afin de protéger les valeurs de la zone :
Des bornes ou des panneaux indiquant l'emplacement et les limites de la zone et précisant clairement les restrictions d'accès devront être placés en des endroits appropriés aux limites de la composante subaérienne de la zone afin d'éviter tout accès par inadvertance.
Les bornes, panneaux et structures érigés à l'intérieur de la zone à des fins scientifiques ou à des fins de gestion seront maintenus en bon état et enlevés lorsqu'ils ne seront plus nécessaires.
Des visites seront effectuées selon les besoins (au moins une fois tous les cinq ans) pour s'assurer que la zone répond toujours aux objectifs pour lesquels elle a été désignée et pour s'assurer que les mesures de gestion et d'entretien sont appropriées.
Une copie du présent plan de gestion devra être mise à disposition dans les installations des quartiers de recherche principaux avoisinant la zone, notamment aux camps du lac Bonney, du lac Hoare, du lac Fryxell, F6 et de New Harbor, ainsi qu'à la station McMurdo et à la base Scott.
Les directeurs des programmes antarctiques nationaux en cours d'exécution dans la région se livreront entre eux à des consultations pour veiller à ce que les dispositions ci-dessus soient mises en œuvre.
4. Durée de désignation
La zone est désignée pour une période indéterminée.
5. Cartes et photographies
Carte 1. - ZSPA n° 172 : Lignes de démarcation de la zone protégée en sous-surface de la partie inférieure du glacier Taylor des Blood Falls. Projection : conique conforme de Lambert ; parallèles types : 1er 77°35'S ; 2e 77°50'S ; méridien central : 161°30'E ; latitude d'origine : 78°00'S ; Datum horizontal et sphéroïde : WGS84 ; équidistance entre les courbes de niveau 200 mètres.
Encart 1 : Situation de la ZGSA n° 2 Vallées sèches de McMurdo dans la région de la mer de Ross.
Encart 2 : Situation du glacier Taylor dans la ZGSA n° 2 Vallées sèches de McMurdo.
Carte 2. - ZSPA n° 172 : Lignes de démarcation de la zone protégée en sous-surface et subaérienne des Blood Falls et site de campement désigné. Projection : conique conforme de Lambert ; parallèles types : 1er 77°43'S ; 2e 77°44'S ; méridien central : 162°16'E ; latitude d'origine : 78°00'S ; Datum horizontal et sphéroïde : WGS84 ; équidistance entre les courbes de niveau 20 mètres.
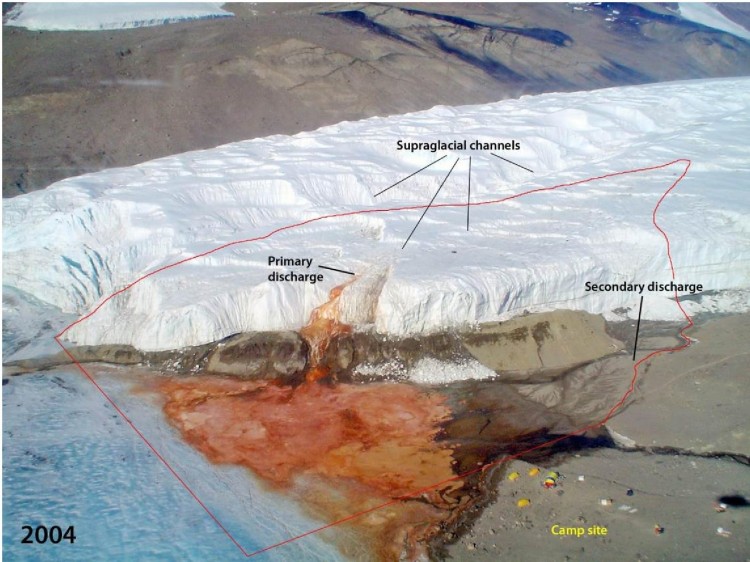
Image 1. Vue aérienne de l'extrémité du glacier Taylor prise en 2004, avec les Blood Falls au centre et le lac Bonney en bas à gauche (Photographe inconnu, 18 novembre 2004). Nota bene que le site de campement est maintenant en grande partie submergé par l'eau du lac Bonney (janvier 2018).
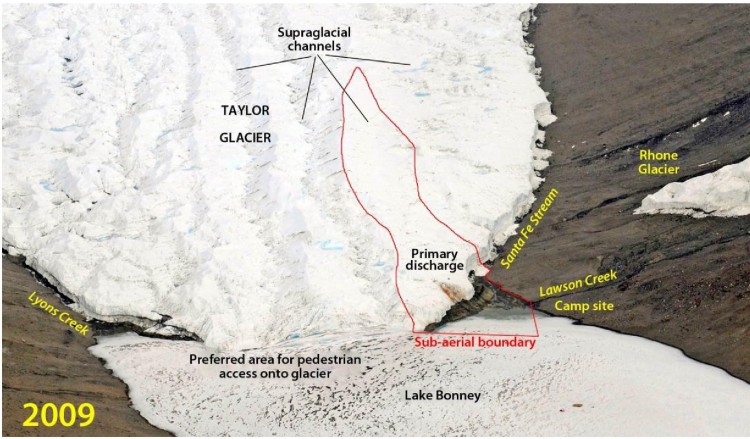
Image 2. Vue aérienne de l'extrémité du glacier Taylor prise en 2009, indiquant l'étendue de la composante subaérienne de la zone. Si l'on compare avec l'image 1, on note combien le débit varie avec le temps (C. Harris, ERA / USAP : 10 décembre 2009).
6. Description de la zone
6 (i) Coordonnées géographiques, bornage et caractéristiques du milieu naturel
Vue d'ensemble
Les Blood Falls (situées à 162°16,288'E, 77°43,329'S) sont un débit d'eau saline riche en fer provenant d'une crevasse située près de l'extrémité du glacier Taylor, dans les vallées sèches de McMurdo, en terre Victoria du Sud. L'eau saumâtre est à l'origine incolore mais, lorsqu'elle coule sur le glacier, elle se transforme en glace blanche contenant des bulles d'air et s'oxyde, produisant la couleur rouge orangé qui lui est particulière. Il reste de nombreuses traces de matière de cette couleur à l'intérieur d'anciennes crevasses et brèches du glacier, en particulier près du site de débit principal. Un débit de surface secondaire, plus petit et moins évident, a été observé à deux reprises (1958 et 1976) à environ 40 mètres au nord du glacier Taylor au bord du delta du cours d'eau Santa Fe (162°16,042'E, 77°43,297'S, carte 2). La composition physique et chimique de ce débit secondaire est comparable à celle de la coulée principale des Blood Falls (Keys 1980).
Le volume et l'étendue physique de la coulée de surface principale des Blood Falls et le volume de glace accumulée qu'elle produit varient dans le temps, allant de quelques centaines à plusieurs milliers de mètres cube de glace saline, et les débits se produisent à des intervalles de un à trois ans, voire plus (Keys 1980). Une proportion inconnue d'eau saumâtre s'écoule dans le lac Bonney avant de geler (par exemple en 1972 et en 1978). A son minimum, le débit se présente comme une petite zone décolorée à l'extrémité du glacier Taylor, mais à son maximum elle peut s'étendre à la surface du lac Bonney sur plusieurs dizaines de mètres (voir images 1 et 2).
Les débits d'eau saumâtre sont d'origine sous-glaciaire, et l'eau de ce débit est de la glace de glacier fondue (Mikucki et al. 2009), mais l'âge et la formation de la source originelle et l'évolution de l'eau saumâtre en sous-surface restent incertains. Les analyses chimiques et isotopiques indiquent qu'un ou plusieurs dépôts de sel marin sont en train de faire fondre et/ou ont fait fondre de la glace du glacier Taylor (Keys 1980). Une analyse approfondie de la topographie sous-glaciaire à environ un à six kilomètres du front glaciaire indique que ce pourrait être l'emplacement du dépôt salin, mais il pourrait aussi y avoir d'autres dépôts plus en amont du glacier. La profondeur et l'étendue de l'eau saumâtre sous-glaciaire à laquelle ils donnent lieu, ainsi que l'emplacement exact et la nature du ou des réservoirs et leur voies d'écoulement, n'ont pas encore été établis de façon définitive (Keys 1980 ; Hubbard et al. 2004).
Limites et coordonnées
Les limites de la zone sont définies de façon à protéger les valeurs du réservoir d'eau saumâtre sous-glaciaire et du débit de surface des Blood Falls et tiennent compte de l'étendue du bassin versant, des connections hydrologiques probables et de l'aspect pratique. Il semble que les connexions hydrologiques et les interactions entre la surface et le lit du glacier Taylor soient limitées. Il n'a donc pas été jugé nécessaire de limiter l'accès à la majorité de la surface du bassin versant et/ou au-dessus de celui-ci. La ligne de démarcation à la surface comprend toutefois une petite zone contenant les débits principal et secondaire confirmés des Blood Falls et inclut une partie de la surface du glacier Taylor s'écoulant directement dans le débit principal, afin d'assurer une protection adéquate aux zones d'écoulement confirmées (carte 2). Les exemples de " sites d'écoulements possibles " indiqués sur la carte 1 ne sont pour le moment pas inclus dans la zone puisqu'ils n'ont toujours pas été confirmés. Ils représentent peut-être les traces de processus de base antérieurs, liés dans le passé au réservoir ou à des caractéristiques qui lui sont associées, plutôt que des sites actuels de débit. Ils ne se déversent d'ailleurs pas dans le réservoir ou dans la coulée principale des Blood Falls.
Par contre, l'étendue des interconnexions sous-glaciaires pourrait être vaste et il s'ensuit que la composante sous-glaciaire visant à protéger la partie principale du bassin versant sous-glaciaire de la partie inférieure du glacier Taylor qui pourrait être connectée avec le réservoir d'eau saumâtre est relativement grande, s'étendant sur environ 50 km vers l'amont du glacier (carte 1). L'étendue de cette zone est considérée comme suffisante pour protéger les valeurs du réservoir, bien qu'il soit reconnu que ces connexions pourraient être plus étendues, puisque le bassin versant s'étend à bonne distance sur le plateau polaire ; la limite occidentale est donc une limite pratique au-delà de laquelle les risques pouvant nuire à la zone sont considérés comme minimes.
En résumé, les limites définissant l'étendue verticale et latérale de la zone visent à :
- protéger l'intégrité du réservoir sous-glaciaire et les zones des débits principal et secondaire confirmés des Blood Falls ;
- tenir compte de l'incertitude vis-à-vis de l'emplacement du réservoir et la connectivité du système hydrologique sous-glaciaire ;
- fournir des lignes de démarcation pratiques correspondant à des bassins versants faciles à cartographier et à identifier sur le terrain ; et
- ne pas imposer de restrictions inutiles aux activités menées à la surface du glacier Taylor et/ou au-dessus.
Les coordonnées principales des limites sont résumées dans le tableau 1.
Tableau 1. - Liste récapitulative des principales coordonnées des limites de la zone protégée (voir cartes 1 et 2).
|
Emplacement |
Désignation |
Longitude (E) |
Latitude (S) |
|
Limites en sous-surface |
|
|
|
|
Débit principal des Blood Falls |
A |
162° 16,305' |
77° 43,325' |
|
Ligne de partage glaciaire des glaciers Taylor et Ferrar, marge sud des collines Kukri |
B |
161° 57,300' |
77° 49,100' |
|
Knobhead, au pied de la crête NE |
C |
161° 44,383' |
77° 52,257' |
|
Vallée Kennar, au centre de la marge du glacier Taylor |
D |
160° 25,998' |
77° 44,547' |
|
Montagne Beehive, au pied de la crête SO |
E |
160° 33,328' |
77° 39,670' |
|
Cirque Mudrey, partie SO |
F |
160° 42,988' |
77° 39,205' |
|
Cirque Mudrey, partie SE |
G |
160° 48,710' |
77° 39,525' |
|
Limites subaériennes |
|
|
|
|
Extrémité du glacier Taylor, affleurement saillant de glace/moraine |
a |
162° 16,639' |
77° 43,356' |
|
Bassin versant supraglaciaire des Blood Falls, partie occidentale |
b |
162° 14,508' |
77° 43,482' |
|
Glacier Taylor, marge nord |
c |
162° 15,758' |
77° 43,320' |
|
Delta du cours d'eau Santa Fe, marge occidentale |
d |
162° 15,792' |
77° 43,315' |
|
Lawson Creek, rocher situé sur la rive ouest |
e |
162° 16,178' |
77° 43,268' |
|
Lac Bonney, ~ 180 mètres à l'est de la rive du delta du cours d'eau Santa Fe |
f |
162° 16,639' |
77° 43,268' |
Sous-surface
La ligne de démarcation en sous-surface comprend la zone d'ablation du glacier Taylor dans son ensemble, allant d'une profondeur de 100 mètres sous la surface jusqu'au lit du glacier. Afin de mieux identifier la limite à la surface, et pour des raisons pratiques liées aux données disponibles quant à la configuration de la ligne de profondeur de 100 mètres au sein du glacier, la marge de surface du glacier Taylor est utilisée pour remplacer cette ligne de profondeur et donc pour déterminer l'étendue latérale de la composante en sous-surface de la zone. La description ci-après définit tout d'abord l'étendue latérale de la composante en sous-surface de la zone, puis son étendue verticale.
La limite de la composante en sous-surface de la zone protégée s'étend depuis le principal site de débit des Blood Falls (162°16,288'E, 77°43,329'S) (identifié par la lettre " A " dans le tableau et sur les cartes 1 et 2) et suit l'extrémité du glacier Taylor vers le sud sur 0,8 km jusqu'à la marge sud du glacier à Lyons Creek. Depuis cet endroit, la limite part en direction sud-ouest sur 19,3 km (carte 1), suivant la marge sud du glacier Taylor jusqu'à l'extrémité occidentale des collines Kukri. Elle continue ensuite vers l'est sur 7,8 km jusqu'à la position approximative de la ligne de partage glaciaire des glaciers Taylor et Ferrar le long de la marge sud des collines Kukri, à 161°57,30'E, 77°49,10'S (" B ", tableau 1, carte 1). La limite se poursuit en direction sud-ouest sur 7,9 km, suivant la ligne approximative du partage glaciaire des glaciers Taylor et Ferrar, jusqu'à l'extrémité orientale de Knobhead, située à 161°44,383'E, 77°52,257'S (" C ", tableau 1, carte 1). La limite suit ensuite la marge sud du glacier Taylor vers l'ouest sur 11,8 km, jusqu'à la ravine Windy, traverse la ravine Windy et continue vers le nord-ouest sur 45,2 km, suivant les marges des glaciers Taylor, Beacon et Turnabout jusqu'à la vallée Kennar, située à 160°25,998'E, 77°44,547'S (" D ", tableau 1, carte 1). La limite part alors en direction nord-est, traversant le glacier Taylor sur 9,5 km jusqu'au pied de la montagne Beehive située à 160°33,328'E, 77°39,670'S (" E ", tableau 1, carte 1). Pour servir de référence visuelle, la limite de la zone protégée est parallèle à une crête distincte visible à la surface du glacier Taylor, immédiatement en aval d'une zone très crevassée.
A partir de la montagne Beehive, la limite se prolonge à l'est sur 5 km jusqu'à la limite entre le cirque Mudrey et le glacier Taylor, à 160°42,988'E, 77°39,205'S (" F ", tableau 1, carte 1). La limite suit alors la bordure du cirque Mudrey sur une longueur de 9,6 km avant de rejoindre le glacier Taylor à 160°48,710'E, 77°39,525'S (" G ", tableau 1, carte 1) et se poursuit ensuite sur 59,6 km en direction du sud-est jusqu'au pied des cascades de glace Cavendish, le long de la marge nord du glacier Taylor. La limite suit alors la marge du glacier Taylor sur 16,9 km en direction nord-est, contournant le lac Simmons et le lac Joyce, puis continue vers l'est sur 15,4 km jusqu'au site de débit principal des Blood Falls (" A ", tableau 1, carte 2).
L'étendue verticale de la composante en sous-surface de la zone se définit en termes de profondeur sous la surface du glacier Taylor (image 3). La limite en sous-surface part d'une profondeur de 100 mètres sous la surface du glacier Taylor jusqu'au lit du glacier, défini comme étant la surface du socle rocheux sur lequel repose le glacier. Le système hydrologique sous-glaciaire, le réservoir d'eau saumâtre des Blood Falls, et toutes les couches contenant un mélange de glace et de sédiments ou des sédiments non consolidés sont inclus dans cette limite. La composante en sous-surface de la zone n'impose aucune restriction supplémentaire aux activités menées à la surface ou dans les premiers 100 mètres à l'intérieur du glacier Taylor.

Image 3. Définition fondée sur des critères de profondeur de l'étendue verticale de la composante en sous-surface de la zone protégée de la partie inférieure du glacier Taylor et des Blood Falls.
Limites subaériennes
La composante subaérienne de la zone comprend le delta du cours d'eau Santa Fe, une partie de l'extrémité occidentale du lac Bonney et un petit bassin versant supraglacial entourant les Blood Falls défini par un système de crêtes de glace persistant dans la morphologie locale du glacier pendant au moins des décennies. La limite sud-est de la composante subaérienne de la zone est marquée par un affleurement saillant de glace et de moraine situé dans le prolongement du front glaciaire du glacier Taylor à 162°16,639'E, 77°43,356'S (identifié par la lettre " a " dans le tableau 1 et sur la carte 2). La limite continue vers le sud-ouest et remonte le glacier sur 900,8 mètres, suivant la marge sud du bassin versant supraglaciaire entourant les Blood Falls jusqu'à la partie la plus à l'ouest du bassin versant supraglaciaire, situé à 162°14,508'E, 77°43,482'S (" b ", tableau 1, carte 2). La limite se poursuit alors vers le nord-est sur une longueur de 594,5 m jusqu'à la marge du glacier Taylor, à 162°15,758'E, 77°43,320'S (" c ", tableau 1, carte 2), le long de la marge septentrionale du bassin versant supraglaciaire. La limite de la zone continue en ligne directe vers le nord-est sur 16,8 mètres, jusqu'à la rive dominant le delta du cours d'eau Santa Fe, à 162°15,792'E, 77°43,315'S (" d ", tableau 1, carte 2). Puis elle longe la rive sur 198,7 mètres en direction nord-est jusqu'à ce qu'elle rejoigne Lawson Creek, à 162°16,178'E, 77°43,268'S (" e ", tableau 1, carte 2). La limite se poursuit ensuite sur 180,5 mètres en ligne droite directement vers l'est jusqu'à un point du lac Bonney situé à 162°16,639'E, 77°43,268'S (" f ", tableau 1, carte 2) avant de continuer en ligne droite plein sud sur 166,5 mètres jusqu'à un affleurement saillant de glace et de moraine.
Climat
Deux stations météorologiques gérées par le programme Long Term Ecological Research (LTER) des vallées sèches de McMurdo sont situées près des Blood Falls ( http://www.mcmlter.org/): La station de " Lac Bonney " (point " a ", 162°27,881'E, 77°42,881'S) situé à ~ 4,5 km à l'est, et celle de " Glacier Taylor " (162°07,881'E, 77°44,401'S), située à ~ 4 km en amont du glacier. La température moyenne annuelle de l'air enregistrée à ces deux stations était d'environ - 17 ºC pendant la période 1993 - 2015. La température la plus basse relevée à ces stations pendant cette période était de - 48,26 ºC, température enregistrée au lac Bonney en août 2008, tandis que la température maximale relevée était de 10,64 ºC, au lac Bonney en décembre 2001. Le mois le plus froid enregistré à ces deux stations était le mois d'août, janvier et décembre étant les mois les plus chauds au lac Bonney et au glacier Taylor respectivement.
La vitesse annuelle moyenne des vents relevée pendant la même période (1993-2015) variait entre 3,89 m/s au lac Bonney et 5,16 m/s au glacier Taylor, avec une vitesse maximale de 44,12 m/s enregistrée au glacier Taylor le 11 mai 2014. La topographie de la vallée Taylor, et notamment la crête de Nussbaum, favorise la formation de systèmes météorologiques isolés dans le bassin du lac Bonney, et limite le flux des vents côtiers dans la région (Fountain et al. 1999).
Les précipitations annuelles moyennes au lac Bonney correspondaient à un équivalent en eau de 340 mm entre 1995 et 2009. Les taux d'ablation du glacier Taylor sont les plus élevés de la région des cascades de glace Cavendish, atteignant un maximum à la base de la ravine Windy (~ 0,4 m a-1), le taux le plus faible étant en amont de la vallée Beacon (~ 0 à 0.125m a-1). Les taux d'ablation de la partie inférieure du glacier Taylor varient habituellement entre 0,15 et 0,3 m a-1 (Bliss et al. 2011).
Géologie et morphologie
La vallée Taylor est composée d'une mosaïque de tills d'âges et de types de rocher divers, notamment : roches de socle précambrien métamorphique (supergroupe de Ross), roches intrusives du Paléozoïque précoce (formation Granite Harbour), une série de roches sédimentaires datant du Dévonien au Jurassique (supergroupe de Beacon) et sills de dolérite de Ferrar datant du Jurassique (Pugh et al. 2003).
On pense que le réservoir sous-glaciaire des Blood Falls est une eau saumâtre d'origine marine provenant d'une incursion marine dans les vallées sèches de McMurdo durant le Pliocène (il y a 3 à 5 Ma BP) ; elle représente peut-être l'eau à l'état liquide la plus ancienne des vallées sèches (Lyons et al. 2005). L'une des théories avancées propose que cette eau saumâtre ait été retenue près de l'extrémité actuelle du glacier Taylor lors du recul des eaux de mer qui s'ensuivit, et qu'elle fut par la suite " scellée " sous le glacier au cours de l'avancée de la glace à la fin du Pliocène ou au Pléistocène (Marchant et al. 1993). On pense que c'est ce dépôt d'eau saumâtre qui forme maintenant un réservoir sous-glaciaire émergeant à la surface de façon épisodique aux sites de débit principal et de débit secondaire latéral. Il a été avancé que l'eau saumâtre s'est modifiée depuis qu'elle a été piégée, notamment du fait des apports liés à l'altération chimique (Keys 1980 ; Lyons et al. 2005 ; Mikucki et al. 2009).
Sols et sédiments
Les sols de la vallée Taylor sont souvent peu développés et se composent en grande partie de sable (95-99 % en poids) (Burkins et al. 2000 ; Barrett et al. 2004). Les concentrations en matière organique des sols de la vallée Taylor sont parmi les plus basses au monde (Campbell & Claridge 1987 ; Burkins et al. 2000) et la teneur en carbone organique des sols du bassin du lac Bonney est particulièrement faible (Barrett et al. 2004). La profondeur des sols de la vallée Taylor varie généralement entre 10 et 30 cm ; en deçà, on y trouve le pergélisol (Campbell & Claridge 1987). En sus des tills glaciaires, le fond de la vallée Taylor est recouvert d'une couche de sédiments lacustres déposés par le glacier autrefois très étendu du lac Washburn, allant jusqu'à environ 300 mètres de profondeur (Hendy et al. 1979 ; Stuiver et al. 1981 ; Hall & Denton 2000).
Les moraines déposées au front du glacier Taylor consistent en des sédiments lacustres transformés datant d'à peu près 300 kA BP (Higgins et al. 2000). Les sédiments que l'on trouve à la marge du glacier Taylor se composent aussi de tills limoneux et sablonneux formés par la fonte de glace basale riche en débris et par l'érosion due aux cours d'eau en bordure de glace (Higgins et al. 2000). Une séquence épaisse de glace basale caractérisée par des sédiments fins qui contiendrait des sels provenant du réservoir sous-glaciaire des Blood Falls a été étudiée grâce à un tunnel creusé dans la marge nord du glacier Taylor (Samyn et al. 2005, 2008 ; Mager 2006 ; Mager et al. 2007). Ces observations suggèrent qu'il existe une interaction entre la base du glacier Taylor et le substrat de sédiment, et il semble qu'il y ait un cycle local de fonte et de gel (Souchez et al. 2004 ; Samyn et al. 2005 ; Mager et al. 2007).
Glaciologie et hydrologie glaciaire
Le glacier Taylor est un glacier émissaire de l'inlandsis oriental de l'Antarctique qui se termine dans le lobe occidental du lac Bonney. Une étude approfondie a récemment été menée pour examiner les dynamiques de la zone d'ablation du glacier Taylor, notamment sa géométrie et le champ de vitesse à la surface (Kavanaugh et al. 2009 a), sa balance de forces (Kavanaugh & Cuffey 2009) et son bilan massique actuel (Fountain et al. 2006 ; Kavanaugh et al. 2009 b). Les résultats indiquent que l'écoulement du glacier est principalement lié à la déformation de la glace froide et que le bilan massique du glacier Taylor est relativement équilibré. Des échantillons de glace provenant de la partie inférieure de la zone d'ablation du glacier Taylor ont été analysés par des chercheurs paléoclimatiques qui les ont datés à la dernière période glaciaire (Aciego et al. 2007). Des études récentes du la partie inférieure du glacier Taylor ont révélé une séquence entière de glace à la structure bien conservée malgré son âge : elle peut dater de 8 à 55 ka BP (Baggenstos et al. 2017), certains pans de glace étant âgés d'au moins 150 ka BP (Severinghaus comm. pers. 2018). Les carottes de glace extraites de cette zone ont servi à analyser les changements dans la composition des gaz atmosphériques (Bauska et al. 2016 ; Petrenko et al. 2017). D'autres programmes récents de recherche glaciologique menés sur le glacier Taylor ont étudié l'évolution des falaises de glace sèches sur le front glaciaire (Pettit et al. 2006 ; Carmichael et al. 2007), mesuré la texture de la glace basale et les gaz qu'elle contient à l'intérieur d'un tunnel sous-glaciaire à proximité du débit principal des Blood Falls (Samyn et al. 2005, 2008 ; Mager et al. 2007) et estimé le bilan énergétique à la surface du glacier (Bliss et al. 2011). Des études de l'hydrologie supraglaciaire du glacier Taylor indiquent que les chenaux de fonte recouvrent environ 40 % de la zone d'ablation de la partie inférieure du glacier Taylor et qu'une proportion importante de la décharge totale dans le lac Bonney provient de la fonte intervenant dans les chenaux (Johnston et al. 2005). Deux grands chenaux traversent la coulée principale des Blood Falls, mais, au vu des températures basses de la glace près de la surface et étant donné que les crevasses ne dépassent pas 100 mètres de profondeur, il semble peu probable qu'il puisse exister des connexions entre les chenaux de fonte à la surface et le réservoir sous-glaciaire des Blood Falls (Cuffey, Fountain, Pettit et Severinghaus, comm. pers. 2010).
L'étendue de l'eau de fonte sous-glaciaire sous le glacier Taylor et ses connexions avec le système des Blood Falls restent pour l'instant incertaines. Les températures basales déduites indiquent que la température de la plus grande partie de la base du glacier Taylor est bien inférieure au point de fusion (Samyn et al. 2005, 2008) et une étude au radar menée par Holt et al. (2006) n'a pas constaté la présence généralisée d'eau liquide sous le glacier Taylor. Les mesures effectuées par Samyn et al. (2005) indiquent une température basale de - 17 °C sur le côté du glacier à proximité des Blood Falls. L'épaisseur de la glace et les gradients de température intraglaciaire probables correspondent toutefois à des températures d'environ - 5 à - 7 °C à la base du glacier situé à 1-3 km des Blood Falls, températures proches de celles qui ont été mesurées dans l'eau saumâtre s'écoulant aux sites primaire et secondaire (Keys 1980). Des études au géoradar indiquent la présence éventuelle d'eau, sans doute hypersaline, dans une dépression du substrat rocheux profonde de 80 mètres et située à une distance de 4 à 6 km de l'extrémité du glacier Taylor (Hubbard et al. 2004).
Le réservoir sous-glaciaire des Blood Falls rejette de l'eau saline de façon épisodique, la plupart du temps par la coulée principale et parfois par le débit latéral secondaire. Des études détaillées menées grâce au VSA (véhicule sous-marin autonome) ENDURANCE (Environmentally Non-Disturbing Under-Ice Robotic Antarctic Explorer) à l'extrémité du glacier Taylor indiquent que l'eau saumâtre sous-glaciaire pourrait se déverser dans le lac Bonney sur presque toute la longueur du front glaciaire (Stone et al. 2010 ; Priscu, pers. comm. 2011). D'autre part, quelques sites ont été identifiés le long des marges nord et sud du glacier Taylor où les couches de glace contiennent des sels et présentent cette même couleur orangée (ces sites sont indiqués comme " sites d'écoulement possibles " sur la carte 1), mais la nature de ces caractéristiques reste encore à définir (Keys 1980 ; Nylen, comm. pers. 2010). Le déclenchement de ces rejets sous-glaciaires est encore mal compris mais il semble que l'eau saumâtre, une fois accumulée sous pression sous le glacier, pourrait se déverser dans un conduit sous-glaciaire discret contrôlant le débit principal. Ce comportement ressemble à certaines débâcles glaciaires apériodiques (jökulhlaups) où les processus de fonte basale et l'évolution des contraintes (par exemple le déplacement physique du glacier Taylor) peuvent créer une brèche dans la retenue de glace basale qui permet à l'eau saumâtre de se déverser, ou bien de chasser le liquide sous-glaciaire de la cuvette rocheuse (Keys 1980 ; Higgins et al. 2000 ; Mikucki 2005). Badgeley et al. (2017) suggère que les Blood Falls font office de " soupape de décharge " pour le système hydrologique où des bassins d'eau saumâtre sous-glaciaire pressurisée en amont des Blood Falls sont injectés de manière intraglaciaire par la formation des crevasses, lui permettant de rester liquide grâce à la cryoconcentration et la libération de chaleur latente. Puis l'eau saumâtre est relâchée, tel un puits artésien épisodique, via les formations de crevasses en surface ayant lieu aux Blood Falls, après avoir été advectée en direction du front glaciaire par le flux de glace.
Le débit principal des Blood Falls est une eau froide (- 6 ºC), riche en carbone organique dissous, en fer et en chlorure de sodium, ayant une conductivité à peu près 2,5 fois supérieure à celle de l'eau de mer (Mikucki et al. 2004 ; Mickuki 2005). Plusieurs sources de données géochimiques appuient la thèse de l'origine marine de la coulée des Blood Falls, dont les caractéristiques sont proches de celles de l'eau de mer. Des études ont prouvé que le volume, l'étendue et la géochimie du débit des Blood Falls varient dans le temps (Black et al. 1965 ; Keys 1979 ; Lyons et al. 2005) et varient aussi selon qu'il s'agisse du flux normal ou d'épisodes de débit rapide (Mikucki 2005).
Ecologie et microbiologie
La coulée des Blood Falls contient une communauté microbienne unique, apparemment d'origine marine (Mikucki & Priscu 2007 ; Mikucki et al. 2009). Les bactéries peuvent sans doute métaboliser les composés ferreux et sulfureux, ce qui leur a permis de survivre dans cet environnement sous-glaciaire très longtemps, peut-être des millions d'années (Mikucki et al. 2009). Les microbes semblent aussi jouer un rôle important dans le cycle du carbone, ce qui a permis à l'écosystème de survivre sans apport externe de carbone (Mikucki & Priscu 2007). Les contrôles primaires des caractéristiques de l'écosystème microbien des Blood Falls fourniront peut-être un analogue des conditions existant sous les calottes de glace polaires de Mars (Mikucki et al. 2004). Un assemblage microbien a été identifié dans des échantillons de glace basale et de sédiments prélevés dans le tunnel creusé dans la marge nord du glacier Taylor (Christner et al. 2010).
L'étude de ces bactéries microbiennes appuie la thèse de l'origine marine du réservoir d'eau saumâtre car les assemblages microbiens des Blood Falls sont analogues à ceux que l'on trouve dans les systèmes marins (Mikucki et al. 2004 ; Mikucki & Priscu 2007). Cet écosystème est maintenant reconnu comme un site important pour les études exobiologiques, notamment comme analogue des masses glaciaires de la planète Mars (Mikucki et al. 2004 ; Mikucki 2005). Il semble que le passé préglaciaire de l'écosystème et du terrain environnant, la lithologie du socle et l'hydrologie du glacier soient les contrôles primaires de l'assemblage microbien des Blood Falls, mais l'ampleur des interactions entre l'écosystème microbien et le système hydrologique glaciaire n'est pas encore bien comprise (Mikucki 2005 ; Mikucki & Priscu 2007).
Les eaux salines sous-glaciaires des Blood Falls rejoignent l'eau comparativement douce de la surface du lac Bonney présente dans la zone entourant le lac dans sa partie occidentale (surnommée la " douve " du lac, car c'est une zone de fonte en été). La zone de la douve est une zone de transition, sa composition géochimique se différenciant de plus en plus des Blood Falls au fur et à mesure que l'on s'éloigne du site de débit principal (Mikucki 2005). Le cours d'eau Santa Fe, alimenté principalement par la fonte de surface du glacier Taylor et coulant le long de sa marge septentrionale, ajoute à la dilution de la coulée des Blood Falls (Mikucki 2005). Lawson Creek se déverse aussi dans la zone et se jette dans le lac Bonney à environ 100 mètres au nord de la coulée principale des Blood Falls.
Les Blood Falls rejettent de façon épisodique de l'eau saline, ainsi que du carbone organique et des bactéries viables, dans le lobe occidental du lac Bonney, entraînant des changements géochimiques et biologiques dans les eaux du lac et lui fournissant des nutriments par ailleurs limités (Lyons et al. 1998, 2002, 2005 ; Mikucki et al. 2004). Des débits ont pu être observés dans le lac Bonney à une profondeur de 20 à 25 mètres, et au-delà de cette profondeur le lac Bonney a une géochimie comparable à celle des Blood Falls, notamment des teneurs en fer élevées et une chimie ionique analogue à celle de l'eau de mer (Black & Bowser 1967 ; Lyons et al. 1998, 2005 ; Mikucki et al. 2004). Certaines études ont montré que les bactéries des zones profondes de l'ouest du lac Bonney sont d'une taille comparable à celle des bactéries des Blood Falls, mais sont beaucoup plus petites que celles que l'on trouve dans les eaux profondes des autres lacs des vallées sèches (Takacs 1999).
Ecologie terrestre
Les communautés d'invertébrés des Blood Falls n'ont pas fait l'objet d'études poussées. Des échantillons de sol, prélevés au bord de la partie occidentale du lac Bonney, indiquent cependant que le Scottnema lindsayae est le nématode le plus abondant dans le bassin du lac Bonney ; ils indiquent aussi la présence d'Eudorylaimus antarcticus et de Plectus antarcticus (Barrett et al. 2004).
Activités humaines et leur impact
Les camps de base dans cette zone ont traditionnellement été situés dans deux aires principales sur la rive nord-ouest du lac Bonney, à proximité de la zone de la douve et de la coulée principale des Blood Falls (carte 2). Le camp contient un certain nombre de sites de tentes marqués par des cercles de pierres. Ceci a causé des perturbations localisées au sol, mais il est peu probable que les activités liées au camp aient eu un impact sur les Blood Falls (Keys, Skidmore, comm. pers. 2010). Jusqu'à il y a peu, il y avait une aire d'atterrissage des hélicoptères à 160 mètres au nord de la coulée principale des Blood Falls et il est pareillement peu probable que son utilisation ait eu des effets nuisibles sur les Blood Falls (Hawes, Skidmore, comm. pers. 2010). Une piste piétonne a été établie à l'ouest de Lawson Creek, parallèle au cours d'eau Santa Fe et au-dessus de celui-ci à environ 50-100 mètres de la marge septentrionale du glacier Taylor. Le trafic piétonnier a rendu la piste très évidente, et elle montre des signes de légère érosion.
Des instruments, notamment un déversoir, avaient été installés par le LTER dans la zone du delta pour la surveillance du cours d'eau Santa Fe (carte 2) ; la majeure partie de ce matériel a été retirée en janvier 2010. Certaines parties du déversoir étaient enchâssées dans les sédiments du cours d'eau et trop difficiles à retirer, ils ont donc été laissées sur place, parce que leur enlèvement aurait causé plus d'impact que de laisser le matériel en place. Du matériel glaciologique hors d'usage a été enlevé de la marge septentrionale du glacier Taylor et de la zone du delta du cours d'eau Santa Fe, mais il se peut que certains éléments aient été laissés dans des endroits inaccessibles et/ou enchâssés dans les sédiments au pied des falaises de glace. Il reste deux tunnels creusés dans la glace basale par des programmes scientifiques antérieurs, le long de la marge septentrionale du glacier Taylor, à ~ 600 mètres et 1 000 mètres des Blood Falls respectivement, qui s'effondreront et fondront avec le temps.
6 (ii) Accès à la zone
Il n'y a pas de restrictions particulières à l'accès au glacier Taylor, aux déplacements sur et/ou au-dessus de sa surface dans la région comprise dans la composante en sous-surface de la zone (Image 3).
L'accès à la composante subaérienne de la zone doit normalement se faire d'abord par hélicoptère sur la zone d'atterrissage désignée, sur la rive nord-ouest du lac Bonney (162°16,47'E, 77°43,17'S, carte 2), et à pied à partir de là. Il est aussi possible d'accéder à la zone en venant à pied depuis le lac Bonney ou de plus haut sur le glacier Taylor.
La voie piétonnière recommandée pour l'accès à la composante subaérienne de la zone depuis la zone d'atterrissage des hélicoptères désignée et le camp est celle qui vient du lac Bonney, évitant autant que possible la coulée glacée d'eau saline colorée et le delta du cours d'eau Santa Fe, remontant l'extrémité du glacier Taylor depuis les pentes situées au sud de la limite de la composante subaérienne (carte 2). Des falaises de glace abruptes présentes le long des marges septentrionales du glacier Taylor empêchent d'accéder à pied à la composante subaérienne de la zone. Plus tard dans la saison, des fosses et des cuvettes qui se forment le long du lac Bonney peuvent empêcher l'accès à la zone.
Une voie piétonnière s'est développée, à une distance d'environ 50 à 100 mètres et parallèle à la marge septentrionale du glacier Taylor, permettant l'accès à la zone plusieurs kilomètres en amont de la zone d'atterrissage des hélicoptères désignée et du camp. Des falaises de glace abruptes présentes le long des marges septentrionales du glacier Taylor empêchent d'accéder à la surface du glacier depuis cet itinéraire.
6 (iii) Emplacement de structures à l'intérieur et à proximité de la zone
Il n'y a pas de structure permanente dans la zone. Deux bornes de surveillance permanentes sont encastrées dans un rocher situé à environ 175 mètres au nord de la zone : le repère géodésique NZAP TP01 est un tube avec filetage femelle (162°16,466'E, 77°43,175'S, élévation 72,7 m) ; le repère géodésique UNAVCO TP02 est un boulon fileté de 5/8" de diamètre (162°16,465'E, 77°43,175'S, élévation 72,8 m). Le rocher se situe dans une zone de terrain en pente sur la rive nord du lac Bonney, à environ 15 m au sud de la zone d'atterrissage des hélicoptères. Un déversoir et une jauge de cours d'eau sont situés à environ 80 m au nord-ouest de la zone à Lawson Creek. Le camp du lac Bonney se situe à environ 4,3 km à l'est de la zone.
6 (iv) Emplacement d'autres zones protégées à proximité
La zone fait partie de la ZGSA n° 2 Vallées sèches de McMurdo. Les zones spécialement protégées de l'Antarctique (ZSPA) les plus proches sont : Glacier Canada (ZSPA n° 131), située à 22 km au nord-est des Blood Falls dans la vallée Taylor ; Terrasse Linnaeus (ZSPA n° 138), située à 31 km au nord-ouest des Blood Falls dans la vallée Wright ; et Vallée Barwick (ZSPA n° 123), située à environ 43 km au nord-ouest des Blood Falls.
6 (v) Zones spéciales à l'intérieur de la zone
Il n'y a aucune aire spéciale à l'intérieur de la zone.
7. Critères de délivrance des permis
7 (i) Conditions générales pour l'obtention d'un permis
L'accès aux composantes subaérienne ou en sous-surface de la zone est interdit sauf si un permis a été délivré à cet effet par les autorités nationales compétentes. Les critères de délivrance d'un permis d'accès à la zone sont les suivants :
- le permis est délivré pour des raisons pédagogiques ou de sensibilisation indispensables qui ne peuvent être satisfaites ailleurs, ou pour des raisons essentielles à la gestion de la zone ;
- les activités autorisées sont conformes au présent plan de gestion ;
- les activités autorisées veilleront, au moyen d'un processus d'évaluation d'impact sur l'environnement, à la protection permanente des valeurs environnementales, écologiques, pédagogiques et scientifiques de la zone ;
- le permis est délivré pour une période limitée ;
- le permis ou une copie certifiée, sera emporté à l'intérieur de la zone.
7 (ii) Accès à la zone et déplacements à l'intérieur de celle-ci
a) Composante en sous-surface (partie inférieure du glacier Taylor) :
L'accès au glacier Taylor, les déplacements sur et/ou au-dessus de sa surface dans la partie de la zone comprenant sa composante en sous-surface ne sont soumis à aucune restriction particulière (image 3).
b) Composante subaérienne (près des Blood Falls).
Accès par voie aérienne et survol
L'atterrissage et le survol de la composante subaérienne de la zone à moins de 100 mètres (328 ft) au-dessus du sol par un aéronef, y compris par de systèmes d'aéronefs pilotés à distance (RPAS), est interdit sauf avis contraire stipulé dans un permis.
Les hélicoptères permettant d'accéder aux Blood Falls devront, de façon générale, éviter les atterrissages dans la composante subaérienne de la zone, et devront de préférence atterrir dans la zone d'atterrissage désignée à cet effet sur la rive nord-ouest du lac Bonney (162°16,47'E, 77°43,17'S, carte 2).
Des hélicoptères ou d'autres aéronefs peuvent être utilisés dans la composante subaérienne de la zone en vue de collecter des données, ou d'effectuer des livraisons de matériel essentiel pour des raisons scientifiques ou de gestion nécessaires pour lesquelles un permis a été délivré, mais ils doivent éviter dans toute la mesure du possible d'emprunter les chenaux supraglaciaires.
Accès en véhicule et utilisation de véhicules dans la zone
Les véhicules sont interdits au sein de la composante subaérienne de la zone.
Accès piéton et déplacements dans la zone
L'accès à la composante subaérienne de la zone et les déplacements à l'intérieur de celle-ci se feront normalement à pied.
Les visiteurs accédant à la composante subaérienne de la zone devront éviter les zones des débits primaire et secondaire des Blood Falls, à moins que les activités autorisées par le permis ne requièrent l'accès à ces sites.
La voie piétonnière recommandée pour l'accès à la composante subaérienne de la zone depuis la zone d'atterrissage des hélicoptères désignée et le camp est celle qui vient du lac Bonney, remontant l'extrémité du glacier Taylor depuis les pentes situées au sud de la limite de la composante subaérienne (carte 2).
Les seuls déplacements au sein de la composante subaérienne de la zone seront ceux nécessaires à la poursuite des activités pour lesquelles un permis a été délivré.
7 (iii) Activités pouvant être conduites à l'intérieur de la zone
Travaux de recherche scientifique qui ne portent pas atteinte aux valeurs de l'écosystème ou scientifiques de la zone et ne nuisent pas à l'intégrité du système des Blood Falls.
Les activités de gestion essentielles, y compris celles de suivi et d'inspection.
Activités à caractère pédagogique (telles que les rapports documentaires, photographiques, audio ou écrits, ou la production de ressources ou services pédagogiques) qu'il n'est pas possible de satisfaire ailleurs ;
Les conditions spécifiques énoncées ci-après s'appliquent aux activités qui sont ou peuvent être menées dans les composantes en sous-surface et subaérienne de la zone :
a) Composante en sous-surface :
Tous les projets nécessitant l'accès à la composante en sous-surface de la zone devront considérer à l'avance les incertitudes quant aux caractéristiques du système hydrologique en sous-surface et le risque que les activités envisagées aient un impact plus que mineur ou transitoire sur les valeurs de la zone. Dans cette perspective, l'évaluation préalable de l'impact sur l'environnement de ces activités devra inclure un examen scientifique détaillé et rigoureux auquel pourront contribuer les experts pertinents.
Ces propositions devront tenir compte du Code de conduite du SCAR sur l'accès à l'environnement aquatique sous-glaciaire et, selon le cas, d'autres protocoles et procédures sur les bonnes pratiques développées afin que l'accès à l'environnement sous-glaciaire se fasse dans des conditions de sûreté et de respect de l'environnement (voir Committee on Principles of Environmental Stewardship for the Exploration and Study of Subglacial Environments 2007 ; Arctic and Antarctic Research Institute 2010 ; Lake Ellsworth Consortium 2011).
Toutes les activités nécessitant l'accès à la composante en sous-surface de la zone devront inclure le suivi de l'efficacité des mesures de contrôle mises en place pour minimiser / prévenir les rejets dans l'environnement.
b) Composante subaérienne :
L'échantillonnage des eaux de fonte des chenaux supraglaciaires se déversant dans la coulée principale des Blood Falls est autorisé, à condition que les mesures appropriées définies en section 7 (vi) soient mises en place afin de réduire au minimum les éventuelles contaminations.
7 (iv) Installation, modification ou enlèvement de structures
Aucune structure ne doit être érigée dans la zone sauf si un permis l'autorise et, à l'exception des bornes et panneaux permanents, toute structure ou installation permanente est interdite.
Toutes les structures, tout le matériel scientifique et tous les repères installés dans la zone doivent être autorisés par un permis et clairement identifiés par pays, nom du principal chercheur et année d'installation. Tous ces éléments doivent être composés de matériaux qui présentent un risque minimum de contamination pour la zone.
L'installation (y compris le choix du site), l'entretien, la modification, ou l'enlèvement de structures doivent s'effectuer d'une façon qui limite les perturbations de l'environnement, ainsi que de la faune et la flore.
Le retrait de matériel spécifique pour lequel le permis est arrivé à expiration relèvera de la responsabilité de l'autorité qui a délivré le permis original et devra figurer dans les critères d'octroi du permis.
Si des équipements sont laissés in situ dans la composante de sous-surface de la zone pendant des périodes prolongées, des dispositions seront prises pour réduire au minimum le risque de contamination et/ou de perte de matériel.
Certains équipements ou matériels pourraient devoir être installés dans des environnements aquatiques sous-glaciaires pour des raisons scientifiques et/ou de suivi (par exemple pour mesurer des processus géophysiques ou biogéochimiques, ou pour surveiller l'impact des activités humaines sur l'environnement sous-glaciaire). Ces installations devront faire l'objet d'un examen spécifique dans l'évaluation de l'impact sur l'environnement de cette activité, et celle-ci inclura aussi la marche à suivre pour leur enlèvement, ainsi que les risques et les bénéfices au cas où leur enlèvement ne serait pas pratique.
7 (v) Emplacement des camps
Les campements sur le glacier Taylor dans la région comprise dans la composante en sous-surface de la zone ne sont sujets à aucune restriction.
Il est interdit de camper dans la composante subaérienne de la zone.
Un camp de base désigné est situé sur la rive nord-ouest du lac Bonney, à environ 150 mètres au nord de la coulée principale des Blood Falls. Il est situé sur une aire rocailleuse en pente douce (vers 162°16,34'E, 77°43,20'S) qui s'étend depuis environ 100 mètres de la rive du lac Bonney et depuis environ 200 mètres au nord-est de Lawson Creek jusqu'à un repère géodésique permanent (TP02) situé à quelque 20 mètres de la rive du lac. Des cercles de pierre marquent les emplacements des tentes. Il convient d'utiliser, dans la mesure du possible, les emplacements de tentes les plus éloignés de la rive du lac Bonney.
7 (vi) Restrictions relatives aux matériaux et organismes pouvant être introduits dans la zone
Aucun animal vivant, aucune forme végétale, aucun micro-organisme et aucun type de sol ne seront introduits délibérément dans la zone et les mesures de précaution décrites ci-dessous seront prises pour la protéger d'une introduction accidentelle.
Les visiteurs devront prendre des précautions spéciales contre toute introduction afin de préserver les valeurs écologiques et scientifiques des Blood Falls et de réduire le risque d'introduction de microbes dans le système des Blood Falls. Sont un motif de préoccupation les introductions d'agents pathogènes, de microbes, d'invertébrés ou de plantes en provenance d'autres sites antarctiques, y compris les stations, ou de régions extérieures à l'Antarctique. Les précautions qui seront prises au sein des composantes subaérienne et en sous-surface de la zone sont les suivantes :
a) Composante en sous-surface :
Pour éviter les introductions de microbes dans la plus grande mesure du possible, tout l'équipement qui sera introduit dans la composante en sous-surface de la zone devra être stérilisé avant d'être utilisé. La stérilisation devra se faire au moyen de méthodes acceptables spécifiées dans l'évaluation d'impact sur l'environnement pour cette activité.
b) Composante subaérienne :
Les visiteurs veilleront à ce que le matériel d'échantillonnage ou les balises soient propres. Les chaussures et autres équipements à utiliser dans la zone (y compris les crampons, les stabilisateurs, les sacs à dos et sacs à provision) devront aussi, dans toute la mesure du possible, être soigneusement nettoyés avant de pénétrer dans la zone. Il est également possible de prendre des chaussures propres (y compris les crampons, etc.) qui ne seront portées qu'à l'intérieur de la zone. Afin de réduire le risque de contamination microbienne, les surfaces exposées des chaussures, le matériel d'échantillonnage et les balises devront être stérilisés avant leur utilisation dans la zone. La stérilisation devra se faire au moyen d'une méthode acceptable comme celle qui consiste à laver dans une solution d'eau contenant 70 % d'éthanol ou dans une solution disponible sur le marché telle que le " Virkon ". Des survêtements de protection stériles devront être portés pour procéder aux échantillonnages dans la partie subaérienne de la zone. Les vêtements protecteurs devront être adaptés pour pouvoir travailler à des températures de - 20 °C ou inférieures et comprendront, au minimum, des combinaisons stériles couvrant bras, jambes et corps et des gants stériles pouvant être enfilés par-dessus les gants classiques. Les protections stériles jetables pour les pieds ne sont pas adaptées aux déplacements sur glacier et ne doivent pas être utilisées.
Aucun herbicide ni pesticide ne doit être introduit dans la zone.
Tout autre produit chimique, y compris les radionucléides ou les isotopes stables, qui peuvent être introduits pour des raisons scientifiques ou raisons de gestion visées dans le permis, seront enlevés de la zone au plus tard à la fin de l'activité pour laquelle le permis a été délivré.
Aucun traceur chimique ne sera introduit dans la composante en sous-surface de la zone, et l'utilisation de traceurs dans la composante subaérienne de la zone se fera conformément à la section " Cours d'eau " des lignes directrices environnementales pour la recherche scientifique figurant à l'appendice B du plan de gestion de la ZGSA n° 2 Vallées sèches de McMurdo.
Il est interdit de déposer dans la zone des combustibles, des aliments et d'autres matériaux à moins qu'ils ne soient nécessaires pour des buts essentiels liés à l'activité pour laquelle le permis a été délivré.
De manière générale, tous les matériaux introduits dans la zone ne le seront que pour une période donnée et ils seront enlevés à la fin de ladite période au plus tard, à moins qu'ils ne soient installés de façon permanente dans l'environnement aquatique sous-glaciaire à des fins scientifiques ou de surveillance, auquel cas les conditions imposées à leur utilisation seront justifiées et détaillées dans l'évaluation de l'impact sur l'environnement pour cette activité.
Tous les matériaux seront entreposés et gérés de telle sorte que les risques posés par leur introduction dans l'environnement soient réduits au minimum.
En cas de déversement susceptible de porter préjudice aux valeurs de la zone, les matériaux seront retirés, dans la mesure où ce retrait n'entraînera pas de conséquences plus graves que de les laisser in situ.
7 (vii) Prélèvement de végétaux, capture d'animaux ou perturbations nuisibles de la faune et la flore
Le prélèvement de végétaux et la capture d'animaux ou les perturbations nuisibles à la faune et la flore indigène sont interdits, sauf sur délivrance d'un permis conforme à l'article 3 de l'annexe II du Protocole au Traité sur l'Antarctique relatif à la protection de l'environnement par l'autorité nationale compétente.
7 (viii) Collecte ou retrait de matériaux non introduits dans la zone par le titulaire du permis
Les matériaux ne peuvent être ramassés ou enlevés de la zone qu'en conformité avec un permis mais ils doivent être limités au minimum requis pour répondre aux besoins scientifiques ou de gestion.
Les débris d'origine humaine qui risquent de porter atteinte aux valeurs de la zone et qui n'ont pas été introduits dans la zone par le détenteur du permis, ou pour lesquels aucune autre autorisation n'a été délivrée, peuvent être enlevés de la zone à moins que l'impact de leur enlèvement ne risque d'être plus grand que si les matériaux étaient laissés in situ. Si tel est le cas, l'autorité compétente doit en être notifiée.
7 (ix) Elimination des déchets
Tous les déchets, y compris les déchets humains, seront enlevés de la zone.
7 (x) Mesures qui peuvent être nécessaires pour continuer de répondre aux objectifs du plan de gestion
Des permis d'accès à la zone peuvent être délivrés pour :
- mener des activités de surveillance et d'inspection dans la zone pouvant comprendre la collecte d'un petit nombre d'échantillons ou de données pour en effectuer l'analyse ou l'examen ;
- installer ou entretenir des panneaux de signalisation, des structures ou des équipements scientifiques ; et
- appliquer des mesures de protection.
7 (xi) Rapports de visite
Les Parties doivent s'assurer que le détenteur principal de chaque permis délivré soumette aux autorités compétentes un rapport décrivant les activités menées dans la zone. Ces rapports doivent inclure les renseignements identifiés dans le formulaire du rapport de visite inclus dans le " Guide pour la préparation des plans de gestion de zones spécialement protégées en Antarctique ". Si nécessaire, l'autorité nationale doit également transmettre une copie du rapport de visite à la Partie étant à l'initiative du plan de gestion, afin de l'aider à la gestion de la zone et dans la révision du plan de gestion.
Les Parties doivent conserver une archive de ces activités et, lors de l'échange annuel d'informations, fournir une description synoptique des activités menées par les personnes relevant de leur juridiction, avec suffisamment de détails pour permettre une évaluation de l'efficacité du plan de gestion. Les Parties doivent, dans la mesure du possible, déposer les originaux ou les copies de ces rapports dans une archive à laquelle le public pourra avoir accès afin de conserver une archive d'usage qui sera utilisée pour toute révision du plan de gestion et pour l'organisation de l'utilisation scientifique de la zone.
Lorsque l'accès à la composante en sous-surface de la zone est requis, les rapports devront également inclure l'emplacement des sites de forage avec une précision de ± 1 mètre et fournir des détails concernant la méthode de forage et le type de fluide de forage employé. Toute contamination de l'environnement de sous-surface fera l'objet d'un rapport. Les rapports feront mention des activités de suivi menées afin de vérifier l'efficacité des mesures mises en place pour contrôler la contamination, notamment celles liées au contrôle microbien.
L'autorité compétente devra être notifiée de toutes les activités entreprises et de toutes les mesures prises ainsi que de tous les matériaux utilisés et non enlevés qui n'étaient pas inclus dans le permis délivré.
8. Bibliographie
Aciego, S.M., Cuffey, K.M., Kvanaugh, J.L., Morse, D.L. & Severinghaus, J.P. 2007. Pleistocene ice and paleo-strain rates at Taylor Glacier, Antarctica. Quaternary Research 68 : 303-13.
Angino, E.E., Armitage, K.B. & Tash, J.C. 1964. Physicochemical limnology of Lake Bonney, Antarctica. Limnology and Oceanography 9 (2) : 207-17.
Arctic and Antarctic Research Institute 2010. Water sampling of the subglacial Lake Vostok. Évaluation globale finale d'impact sur l'environnement. Expédition Antarctique de Russie, Institut de recherche d'Arctique et d'Antarctique. Saint-Pétersbourg, Russie.
Badgeley, J.A., Pettit, E.C., Carr, C.G., Tulaczyk, S., Mikucki, J.A., Lyons, W.B. & équipe scientifique du MIDGE 2017. An englacial hydrologic system of brine within a cold glacier : Blood Falls, McMurdo Dry Valleys, Antarctica. Journal of Glaciology 63(239) : 387-400.
Baggenstos, D., Bauska, T.K., Severinghaus, J.P., Lee, J.E., Schaefer, H., Buizert, C., Brook, E.J., Shackleton, S. & Petrenko, V.V. 2017. Atmospheric gas records from Taylor Glacier, Antarctica, reveal ancient ice with ages spanning the entire last glacial cycle. Climate of the Past 13(7) : 943-58. https://doi.org/10.5194/cp-13-943-2017, 2017.
Barrett, J.E., Virginia, R.A., Wall, D.H., Parsons, A.N., Powers, L.E. & Burkins, M. B. 2004. Variation in biogeochemistry and soil biodiversity across spatial scales in a polar desert ecosystem. Ecology 85 (11) : 3105-18.
Bauska, T.K., Baggenstos, D., Brook, E.J., Mix, A.C., Marcott, S.A., Petrenko, V.V., Schaefer, H., Severinghaus J.P. & Lee J.E. 2016. Carbon isotopes characterize rapid changes in atmospheric carbon dioxide during the last deglaciation. PNAS 113(13) : 3465-70.
Black, R.F. & Bowser, C.J. 1967. Salts and associated phenomena of the termini of the Hobbs and Taylor Glaciers, Victoria Land, Antarctica. International Union of Geodesy and Geophysics, Commission on Snow and Ice. Publication 79 : 226-38.
Black, R. F., Jackson, M. L. & Berg, T. E., 1965. Saline discharge from Taylor Glacier, Victoria Land, Antarctica. Journal of Geology 74 : 175-81.
Bliss, A.K., Cuffey, K.M. & Kavanaugh, J.L. 2011. Sublimation and surface energy budget of Taylor Glacier, Antarctica. Journal of Glaciology 57 (204) : 684-96.
Burkins, M. B., Virginia, R.A., Chamberlain, C.P. & Wall, D.H. 2000. Origin and Distribution of Soil Organic Matter in Taylor Valley, Antarctica. Ecology 81 (9) : 2377-91.
Campbell, I.B. & Claridge, G.G.C. 1987. Antarctica : soils, weathering processes and environment (Développements dans Soil Science 16). Elsevier, New York.
Carmichael, J.D., Pettit, E.C., Creager, K.C. & Hallet, B. 2007. Calving of Taylor Glacier, Dry Valleys, Antarctica. Eos Transactions AGU 88 (52), Suppléments à la réunion d'automne, Résumé C41A-0037.
Christner, B.C., Doyle, S.M., Montross, S.N., Skidmore, M. L., Samyn, D., Lorrain, R., Tison, J. & Fitzsimons, S. 2010. A subzero microbial habitat in the basal ice of an Antarctic glacier. Réunion d'automne de l'AGU 2010, Résumé B21F-04.
Committee on the Principles of Environmental Stewardship for the Exploration and Study of Subglacial Environments, 2007. Exploration of Antarctic Subglacial Aquatic Environments : Environmental and Scientific Stewardship. Polar Research Board, National Research Council, National Academies Press, Washington D.C. ( http://www.nap.edu/catalog/11886.html?).
Foley, N., Tulaczyk, S., Auken, E., Schamper, C., Dugan, H., Mikucki, J., Virginia, R. & Doran, P. 2015. Helicopter-borne transient electromagnetics in high-latitude environments : An application in the McMurdo Dry Valleys, Antarctica. Geophysics 81(1) : WA87-WA99.
Fountain, A.G., Lyons, W.B., Burkins, M. B. Dana, G.L., Doran, P.T., Lewis, K.J., McKnight, D.M.,
Moorhead, D.L., Parsons, A.N., Priscu, J.C., Wall, D.H., Wharton, Jr., R.A. & Virginia, R.A. 1999. Physical controls on the Taylor Valley ecosystem, Antarctica. BioScience 49 (12) : 961-71.
Fountain, A.G., Nylen, T.H., MacClune, K.J., & Dana, G.L. 2006. Glacier mass balances (1993-2001) Taylor Valley, McMurdo Dry Valleys, Antarctica. Journal of Glaciology 52 (177) : 451-465.
Lake Ellsworth Consortium 2011. Proposed exploration of subglacial Lake Ellsworth, Antarctica. Projet d'évaluation globale d'impact sur l'environnement. British Antarctic Survey, Cambridge.
Hall, B.L. & Denton, G.H. 2000. Radiocarbon Chronology of Ross Sea Drift, Eastern Taylor Valley, Antarctica : Evidence for a Grounded Ice Sheet in the Ross Sea at the Last Glacial Maximum. Geografiska Annaler : Series A, Physical Geography 82 (2-3) : 305-36.
Hamilton, W. L., Frost, I. C. & Hayes P. T. 1962. Saline Features of a Small Ice Platform in Taylor Valley, Antarctica. USGS Professional Paper 450B. US Geological Survey : B73-76.
Hendy, C.H., Healy, T.R., Rayner, E.M., Shaw, J. & Wilson, A.T. 1979. Late Pleistocene glacial chronology of the Taylor Valley, Antarctica, and the global climate. Quaternary Research 11 (2) : 172-84.
Higgins, S.M., Denton, G. H. & Hendy, C. H. 2000. Glacial Geomorphology of Bonney Drift, Taylor Valley, Antarctica. Geografiska Annaler. Series A, Physical Geography 82 (2-3) : 365-89.
Holt, J.W., Peters, M. E., Morse, D.L., Blankenship, D.D., Lindzey, L.E., Kavanaugh, J.L. & Cuffey, K.M. 2006. Identifying and characterising subsurface echoes in airborne radar sounding from a high-clutter environment in the Taylor Valley, Antarctica. 11e Conférence internationale sur les géoradars, 19-22 juin 2006, Columbus Ohio.
Hubbard, A., Lawson, W., Anderson, B., Hubbard, B. & Blatter, H. 2004. Evidence of subglacial ponding across Taylor Glacier, Dry Valleys, Antarctica. Annals of Glaciology 39 : 79-84.
Johnston, R.R., Fountain, A.G. & Nylen, T.H. 2005. The origins of channels on lower Taylor Glacier, McMurdo Dry Valleys, Antarctica, and their implication for water runoff. Annals of Glaciology 40 : 1-7.
Kavanaugh. J.L. & Cuffey, K.M. 2009. Dynamics and mass balance of Taylor Glacier, Antarctica : 2. Force balance and longitudinal coupling. Journal of Geophysical Research 114 : F04011.
Kavanaugh. J.L., Cuffey, K.M., Morse, D.L., Conway, H. & Rignot, E. 2009a. Dynamics and mass balance
of Taylor Glacier, Antarctica : 1. Geometry and surface velocities. Journal of Geophysical Research 114 : F04010.
Kavanaugh. J.L., Cuffey, K.M., Morse, D.L., Bliss, A.K. & Aciego, S.M. 2009b. Dynamics and mass balance of Taylor Glacier, Antarctica : 3. State of mass balance. Journal of Geophysical Research 114 : F04012.
Keys, J.R. 1979. The saline discharge at the terminus of Taylor Glacier. Antarctic Journal of the United States 14 : 82-85.
Keys, J.R 1980. Salts and their distribution in the McMurdo region, Antarctica. Chapitre 8 d'une thèse de doctorat non publiée soutenue à l'université de Victoria à Wellington NZ, et centre de recherche polaire Byrd, Columbus, Ohio : 240-82.
Lyons, W.B., Nezat, C.A., Benson, L.V., Bullen, T.D., Graham, E.Y., Kidd, J., Welch, K.A. & Thomas, J.M. 2002. Strontium isotopic signatures of the streams and lakes of Taylor Valley, Southern Victoria Land, Antarctica : chemical weathering in a polar climate. Aquatic Geochemistry 8 (2) : 75-95.
Lyons, W.B. Tyler, S.W. Wharton Jr R.A., McKnight D.M. and Vaughn B.H. 1998. A Late Holocene desiccation of Lake Hoare and Lake Fryxell, McMurdo Dry Valleys, Antarctica. Antarctic Science 10 (3) : 247-56.
Lyons, W.B., Welch, K.A., Snyder, G., Olesik, J., Graham, E.Y., Marion, G.M. & Poreda, R.J. 2005. Halogen geochemistry of the McMurdo dry valleys lakes, Antarctica : Clues to the origin of solutes and lake evolution. Geochimica et Cosmochimica Acta, 69 (2) : 305-23.
Mager, S., Fitzsimons, S., Frew, R. & Samyn, D. 2007. Stable isotope composition of the basal ice from Taylor Glacier, southern Victoria Land, Antarctica. Géological Survey (Etats-Unis) et les National Academies ; USGS OF-2007-1047, Résumé détaillé 109.
Mager, S. 2006. A compositional approach to understanding the formation of basal ice in Antarctic glaciers. Thèse de doctorat non publiée ; Université d'Otago, Dune din, Nouvelle-Zélande.
Marchant, D. R., Denton, G. H. & Sugden, D. E. 1993. Miocene glacial stratigraphy and landscape evolution in the western Asgard Range, Antarctica. Geografiska Annaler 75 : 269-302.
Mikucki, J. A. 2005. Microbial Ecology of an Antarctic Subglacial Environment. Thèse de doctorat non publiée, Université d'État du Montana, Bozeman, Montana.
Mikucki, J.A., Foreman, C.M., Sattler, B., Lyons, W.B. & Priscu, J.C. 2004. Geomicrobiology of Blood Falls : An iron-rich saline discharge at the terminus of the Taylor Glacier, Antarctica. Aquatic Geochemistry 10 : 199-220.
Mikucki, J.A., Pearson, A., Johnston, D.T. Turchyn, A.V., Farquhar, J., Schrag, D.P., Anbar, A.D., Priscu, J.C. & Lee, P.A. 2009. A contemporary microbially maintained subglacial ferrous “ocean”. Science 324 : 397-400.
Mikucki, J.A. & Priscu, J.C. 2007. Bacterial diversity associated with Blood Falls, a subglacial outflow from the Taylor Glacier, Antarctica. Applied and Environmental Microbiology 73 (12) : 4029-39.
Mikucki, J.A., Auken, E., Tulaczyk, S., Virginia, R.A., Schamper, C., Sørensen, K.I., Doran, P.T., Dugan, H. & Foley, N. 2015. Deep groundwater and potential subsurface habitats beneath an Antarctic dry valley. Nature Communications 6 : 6831.
Petrenko, V.V., Smith, A.M., Schaefer, H., Riedel, K., Brook, E., Baggenstos, D., Harth, C., Hua, Q., Buizert, C., Schilt, A., Fain, X., Mitchell, L., Bauska, T.K., Orsi, A., Weiss, R.F. & Severinghaus, J.P. 2017. Minimal geologic methane emissions during Younger Dryas - Preboreal abrupt warming event. Nature 548 : 443-46.
Pettit, E.C., Nylen, T.H., Fountain, A.G. & Hallet, B. 2006. Ice Cliffs and the Terminus Dynamics of Polar Glaciers. Eos Transactions AGU 87 (52) Suppléments à la réunion d'automne, Résumé C41A-0312.
Pugh, H.E., Welch, K.A., Lyons, W.B., Priscu, J.C. & McKnight, D.M. 2003. The biogeochemistry of Si in the McMurdo Dry Valley lakes, Antarctica. International Journal of Astrobiology 1 (4) : 401-13.
Samyn, D., Fitzsimmons, S.J. & Lorrain, R.D. 2005. Strain-induced phase changes within cold basal ice from Taylor Glacier, Antarctica, indicated by textural and gas analyses. Journal of Glaciology 51 (175) : 165-69.
Samyn, D., Svensson, A. & Fitzsimons, S. 2008. Discontinuous recrystallization in cold basal ice from an Antarctic glacier : dynamic implications. Journal of Geophysical Research 113 F03S90, doi : 101029/2006JF000600.
SCAR 2011. Code de conduite du SCAR pour l'exploration et la recherche dans des environnements aquatiques sous-glaciaires. Document d'information 33 (IP 33), XXXXIVe RCTA, Buenos Aires.
Souchez, R., Samyn, D., Lorrain, R., Pattyn, F. & Fitzsimons, S. 2004. An isotopic model for basal freeze-on associated with subglacial upward flow of pore water. Geophysical Research Letters 31 L02401.
Spigel, R.H., Priscu, J.C., Obryk, M. K., Stone, W. & Doran, P.T. (sous presse 2018). The physical limnology of a permanently ice-covered and chemically stratified Antarctic lake using high resolution spatial data from an autonomous underwater vehicle. Limnology and Oceanography.
Stone, W., Hogan, B., Flesher, C., Gulati, S., Richmond, K., Murarka, A., Kuhlman, G., Sridharan, M. , Siegel, V., Price, R.M., Doran, P.T. & Priscu, J. 2010. Design and Deployment of a Four-Degrees-of-Freedom Hovering Autonomous Underwater Vehicle for sub-Ice Exploration and Mapping. Proceedings of the Institution of Mechanical Engineers, Part M : Journal of Engineering for the Maritime Environment 224 : 341-61.
Stuvier, M. , Denton, G.H., Hughes, T.J. & Fastook, J.L. 1981. History of the marine ice sheet in West Antarctica during the last glaciation : a working hypothesis. Dans Denton, G. H. & Hughes, T. H., Eds. The last great ice sheets. Wiley-Interscience, New York : 319-436.
Takacs, C.D. 1999. Temporal Change in Bacterial Plankton in the McMurdo Dry Valleys. Thèse de doctorat non publiée ; Université d'Etat du Montana, Bozeman, Montana.
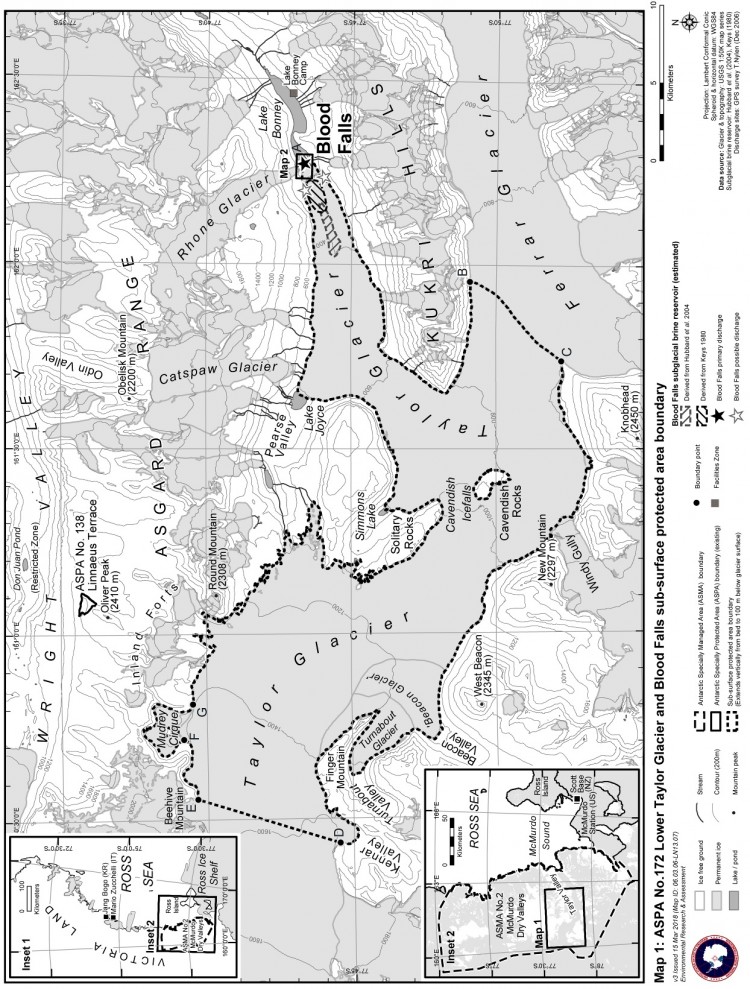
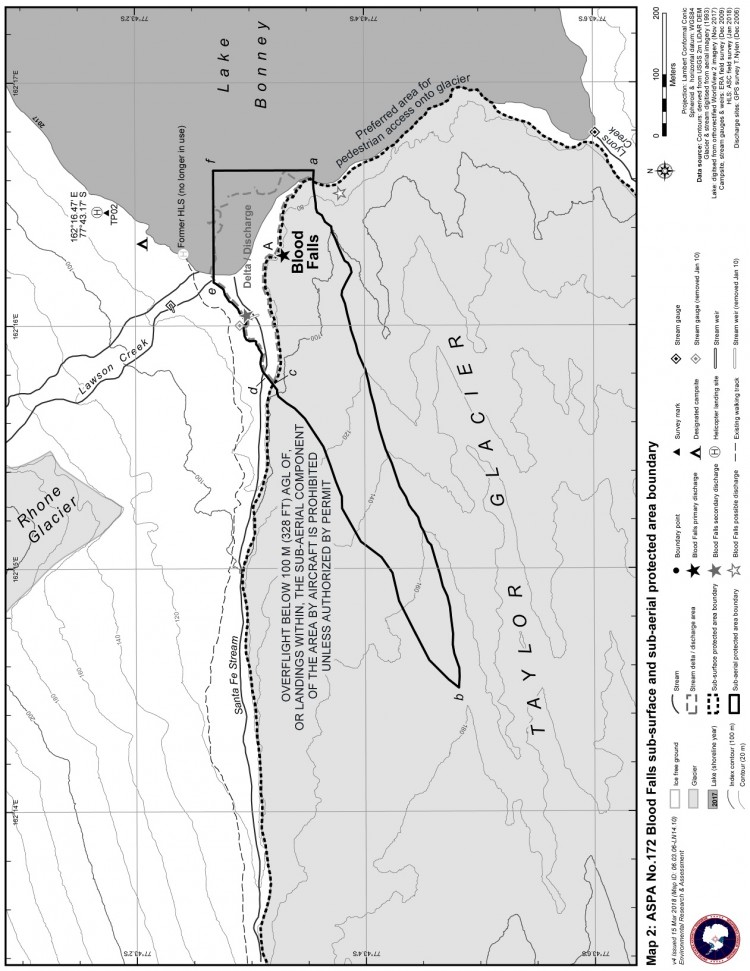
(1) Entrée en vigueur : 16 août 2018.